
The Role of Trees in Agroecology and Sustainable Agriculture in the Tropics
This chapter was previously published in Leakey, R.R.B., 2014. Annual Review of Phytopathology 52, 113–133, with permission of Annual Reviews
Abstract
In conventional modern agriculture trees are typically seen as a hindrance to intensive agriculture and are thus cleared from fields with the subsequent loss of the biodiversity and its role in a very diverse set of agroecological functions. This deforestation leads to the loss of soil fertility, land degradation, declining crop yields and loss of livelihoods, as well as greenhouse gas emissions to the atmosphere and threats to the survival of wildlife. This chapter examines how all these impacts can be reversed by integrating trees back into farmland and developing an alternative model for farm intensification – one especially appropriate for smallholder farming systems in the tropics.
Keywords
Agroecosystem; agroforestry; biodiversity; cocoa; coffee; conservation; ecological succession; Nelder fan; pests and diseases; Replacement series; wildlife
Summary
Shifting agriculture in the tropics has been replaced by sedentary smallholder farming on a few hectares of degraded land. To address both low yields and low income, the soil fertility, the agroecosystem functions, and the source of income can be restored by diversification with nitrogen-fixing trees and the cultivation of indigenous tree species that produce nutritious and marketable products. Biodiversity conservation studies indicate that mature cash crop systems, such as cacao and coffee with shade trees, provide wildlife habitat that supports natural predators, which in turn reduce the numbers of herbivores and pathogens. This review offers suggestions on how to examine these agroecological processes in more detail for the most effective rehabilitation of degraded land. Evidence from agroforestry indicates that in this way productive and environmentally friendly farming systems that provide food and nutritional security, as well as poverty alleviation, can be achieved in harmony with wildlife.
Introduction
The future of agriculture is the focus of much international debate (Cribb, 2010; Garnett et al., 2013; Giovannucci et al., 2012; IAASTD, 2009). The main area of consensus is that it needs to be more sustainable and more productive, and that business as usual is not an option (GEO, 2007; Hassan et al., 2005; IAASTD, 2009; Molden, 2007; Royal Society, 2009). The problems are numerous and complex, especially in the tropics and subtropics, where interactions between both biophysical and socioeconomic issues are affecting agricultural productivity, mainly through a combination of land degradation driven by population growth and a declining resource of productive land for agriculture. This has led to farmers becoming sedentary smallholders without the financial resources to replenish soil fertility (Leakey, 2012b, 2013).
In his book The Coming Famine, Julian Cribb (2010) says that “… the challenge facing the world’s 1.8 billion men and women who grow our food is to double their output of food—using far less water, less land, less energy and less fertilizer. They must accomplish this … amid more red tape, economic disincentives, and corrupted markets, and in the teeth of spreading drought.” Adding more detail to this scenario, the International Food Policy Research Institute in Washington has indicated that the world population will rise from the current 7 billion to approximately 9 billion by 2050, and that to support all these people, food production will have to increase by 70% (IFPRI, 2011). However, it is not clear whether this projection is accepting the status quo in terms of the living standards of the poor in least-developed countries or of the effect on the environment. The real need is, of course, to feed marginalized people and also to improve their lives.
These projections make pretty gloomy reading, but is it a lost cause? The context of the problems here is important. We need to understand that hundreds of millions of poor farmers are trying to support their families on a small area of land without access to fertilizers and other technologies. This new sedentary lifestyle is the result of population growth and the lack of new forest to clear for the practice of shifting cultivation. As sedentary farmers, they have to survive, feed, and provide for all the daily needs of their families on approximately two to five hectares of cleared land without a source of income or the lifeline of social services if something goes wrong.
Regarding solutions, much of the focus of discussion is polarized between whether to improve productivity through biotechnology and genetic modification or through better soil husbandry and organic farming. A third commonly advocated solution is to pay greater attention to agroecology, which is certainly relevant to addressing the loss of biodiversity and ecosystem function. However, achieving a sustainable future for agriculture will involve more than just improving agroecological functions. Instead, it is necessary to take a more holistic view of a problem that encompasses the need for improved soil fertility and health; reduced risk of pest, disease, and weed outbreaks; improved crop yields; improved rural and urban livelihoods; and opportunities for economic growth.
Understanding the Problem Before Seeking Solutions
Current estimates are that land degradation affects 2 billion hectares (38% of world cropping area) (Eswaran et al., 2006), with soil erosion affecting 83% of the global degraded area (Bai et al., 2008). The problems are especially severe in Africa, where more than 80% of countries are nitrogen-deficient, and where nutrient loss has been estimated at 9–58 kg ha−1 year−1 in the 28 worst affected countries and 61–88 kg ha−1 year−1 in 21 others (Chianu et al., 2012). Nitrogen (N), phosphorus (P), potassium (K) deficiencies occur on 59%, 85%, and 90% of harvested land area, respectively (Chianu et al., 2012).
Understanding the complexity of the numerous and interacting socioeconomic and biophysical causal factors underlying land degradation and the problems of modern agriculture in the tropics and subtropics is difficult. In the past, they have been lumped together as a downward spiral in which poverty drives land degradation and land degradation drives further poverty, but this is a gross simplification (Scherr, 2000). In an attempt to make the concept more meaningful, Leakey (2010) added some steps to a conceptual diagram of the land degradation and social deprivation cycle. In this highly interactive cycle, a desire for security and wealth drives deforestation, overgrazing, and unsustainable use of soils and water, all of which cause agroecosystem degradation. In farmers’ fields, this is seen as soil erosion, breakdown of nutrient cycling, and the loss of soil fertility and structure. The consequence of this degradation is the loss of biodiversity and the breakdown of ecosystem functions that regulate food chains, nutrient cycling, and the incidence and severity of pests, diseases, and weeds. All of these things result in lower crop yields, which in turn lead to hunger, malnutrition, and increased health risks, all of which manifest as declining livelihoods and so return the cycle to a desire for security and wealth. It is recognized that at all of the steps within this conceptual diagram, there are a range of socioeconomic and biophysical influences that will determine the speed of the downward progress at any particular site. Such factors include access to markets, land tenure, and local governance; external factors, such as natural disasters and conflict and war; and economic drivers, such as international policy and trade agreements.
The net result highlighted by this conceptual cycle of land degradation and social deprivation is that in developing countries where farmers are poor and must rely on a small piece of land for all their household needs, the benefits expected from existing agricultural technologies, such as improved crop varieties and livestock breeds, are constrained by a need for income to purchase the inputs essential for food production. As a consequence, crop yields decline as the soil fertility is depleted. This leads to what is called the yield gap in food crops, which is the difference between the yield potential of new modern varieties and the yield actually achieved by a farmer (Tittonell and Giller, 2013).
Addressing Soil Fertility
Conventional agriculture would advocate that inorganic fertilizers be used to make up these deficiencies, but unfortunately in many of the least-developed countries where this need is greatest, the farmers are too poor to purchase them. Even when available, they are typically much more expensive than in industrialized countries. An alternative promoted by the organic agriculture movement is the use of manure, compost, and mulching. The problem here is that organic manure is seldom available in the quantities (10–40 Mg ha−1 year−1) needed for large areas (Mafongoya et al., 2006).
Another alternative is to utilize biological nitrogen-fixing technologies, such as 2-year improved fallows, relay cropping, or Evergreen Agriculture, by which nitrogen-deficient soils are enriched with fast-growing leguminous trees, shrubs, or vines. A study involving 16 tree species found that the rate of nitrogen fixation was higher than the 23–176 kg N ha−1 year−1 for food or fodder legumes (Herridge et al., 2008), and in high-density plantings, such as improved fallows or fodder banks, could be as high as 300–650 kg N ha−1 year−1 (Nygren et al., 2012). Together, these research findings on crop production conform to the general experience of adopting farmers who report yield increases of two- to threefold. The combination of inorganic fertilizers and biological nitrogen can, however, be synergistic.
Agroforestry has a long and well-documented history in the use of low-cost, biological nitrogen fixation by leguminous trees and shrubs to restore nitrogen deficiencies in the soils across Africa and other areas of the tropics and subtropics (Sanchez, 1976; Young, 1997). A recent meta-analysis of 94 studies in sub-Saharan Africa found that these tree technologies increased maize yields by 0.7–2.5 Mg ha−1 (or 89–318%) (Sileshi et al., 2008a). Furthermore, these yield increases were more stable than monocultures treated wih recommended applications of artificial fertilizer (Sileshi et al., 2011). Indirectly, leguminous trees can also recycle other macro- and micronutrients through uptake from their more extensive and deeper root systems (which act as a safety net to absorb deep nutrients) and subsequent leaf litter fall and fine root turnover (Mweta et al., 2007). By comparison with artificial fertilizers, nitrogen of tree origin is less likely to be lost to, or pollute, groundwater.
Environmentally, biological nitrogen fixation by leguminous trees and shrubs is important for soil health, as it increases the organic matter and carbon contents of soil, which improve soil structure (aggregate stability, porosity, and hydraulic conductivity), reduce soil erosion, and promote greater water infiltration. Soil organic matter content is important in all agroecosystems, but it is especially important in areas where rainfall can be a limiting factor. Ecologically, the organic nitrogen not taken up by the crops is stored in the soil organic matter pools (Schroth et al., 2001) and so is available to other organisms, such as mycorrhizal fungi and earthworms (Sileshi et al., 2008b,c). This nitrogen resource is less likely to be leached and to pollute groundwater than is the use of inorganic fertilizers. Leguminous trees have also been shown to considerably improve rain-use efficiency (the ratio of aboveground net primary production to annual rainfall) (Sileshi et al., 2011) Likewise, water-use efficiency was higher in maize intercropped with leguminous trees than in the sole maize (Chirwa et al., 2007). In general, the use of biological nitrogen fixation to improve soil fertility also leads to crops less susceptible to pests and diseases (Schroth et al., 2000). On the negative side, as with inorganic fertilizers, there can be N2O emissions associated with higher levels of soil nitrogen.
Biodiversity and Agroecological Functions
The role of ecological processes in agricultural sustainability has been studied for many years (Swift and Anderson, 1994), and its importance for the future of global agriculture is well recognized (Tscharntke et al., 2012). However, its application has not occurred on a large-enough scale to have impact on the global problem of land degradation. The challenge, therefore, is to demonstrate conclusively and to the satisfaction of policy makers, agribusiness, and academics that diverse mixed cropping systems can be productive while using natural resources sustainably. This concept is described as sustainable intensification (Garnett et al., 2013). Care, of course, has to be taken not to introduce species that cohost serious pests and diseases of the planted crops (Schroth et al., 2000).
Previously we saw that integrating leguminous trees and shrubs in cropping systems can initiate the process of rebuilding soil fertility and also boost agroecosystem functions. In recognition of this role of trees, Leakey (Leakey, 1996, 1999b) described agroforestry practices as “phases in the development of a productive agroecosystem, akin to the normal dynamics of natural ecosystems … and the passage toward a mature agroforest of increasing ecological integrity. By the same token, with increasing scale, the integration of various agroforestry practices into the landscape is like the formation of a complex mosaic of patches in an ecosystem, each of which is composed of many niches … occupied by different organisms, making the system ecologically stable and biologically diverse.” Over recent years, there has been a considerable increase in the number of studies examining the biological diversity associated with complex multistrata agroforestry systems, especially cocoa (Clough et al., 2009a) and coffee (Somarriba et al., 2004), and their comparison with other farming systems (Collins and Qualset, 1999; Schroth et al., 2004; Table 2.1). Many of these studies find that agroforestry systems maintain a level of biodiversity that is considerably higher than other agricultural systems but generally a little lower than that of natural forest. Again, a typical finding is that agroforests provide habitat suitable for forest-dependent species and so, in most cases, are important for wildlife conservation.
Table 2.1
Published assessments of biodiversity in agroforestry systems (see also Moguel and Toledo, 1999; Perfecto et al., 1996).
| Organisms | Country | Crop | Reference |
| Phytotelmata plants and ceratopogonid pollinators | Brazil | Cocoa | Fish and Soria (1978) |
| Bat and bird | Brazil | Cocoa | Faria et al. (2006) |
| Bat | Brazil | Cocoa | Faria and Baumgarten (2007) |
| Ants | Brazil | Cocoa | Delabie et al. (2007) |
| Soil and litter fauna | Brazil | Cocoa | da Silva Moço et al. (2009) |
| Sloth | Brazil | Cocoa | Cassano et al. (2011) |
| Plants | Cameroon | Cocoa | Sonwa et al. (2007) |
| Soil fauna | Cameroon | Cocoa | Norgrove et al. (2009) |
| Mammals | Cameroon | Cocoa | Arlet and Molleman (2010) |
| Birds | Costa Rica | Cocoa | Reitsma et al. (2001) |
| Dung beetles, mammals (including bats), birds | Costa Rica | Cocoa and banana | Harvey et al. (2006a,b), Harvey and González Villalobos (2007) |
| Sloth | Costa Rica | Cocoa | Vaughan et al. (2007) |
| Epiphytes | Ecuador | Cocoa | Andersson and Gradstein (2005), Haro-Carrión et al. (2009) |
| Birds | Indonesia | Cocoa | Abrahamczyk et al. (2008) |
| Epiphytes | Indonesia | Cocoa | Sporn et al. (2009) |
| Amphibians, reptiles, and ants | Indonesia | Cocoa | Wanger et al. (2009, 2010a,b) |
| Ants | Indonesia | Cocoa | Wielgoss et al. (2010) |
| Rats | Indonesia | Cocoa | Weist et al. (2010) |
| Monkeys | Mexico | Cocoa | Muñoz et al. (2006) |
| Birds | Mexico | Cocoa | Greenberg (2000) |
| Homoptera | Costa Rica | Coffee | Rojas et al. (2001a,b) |
| Birds | Dominican Republic | Coffee | Wunderle and Latte (1998) |
| Birds and other seed dispersal agents | Ecuador | Coffee | Lozada et al. (2007) |
| Mammals | India | Coffee | Bali et al. (2007) |
| Termites | India | Coffee | Gowda et al. (1995) |
| Soil coleoptera | Mexico | Coffee | Nestel et al. (1993) |
| Mammals | Mexico | Coffee | Gallina et al. (1996) |
| Ants | Mexico | Coffee | Perfecto and Snelling (1995) |
| Arthropod | Mexico | Coffee | Perfecto et al. (1997) |
| Birds | Mexico | Coffee | Perfecto et al. (2004) |
| Birds | Mexico | Coffee | Greenberg et al. (1997) |
| Ants and birds | Panama | Coffee | Roberts et al. (2000a) |
| Ants | Panama | Coffee | Roberts et al. (2000b) |
| Collembola | Indonesia | Rubber | Deharveng (1992) |
| Mammals | Indonesia | Rubber, damar, and durian | Sibuea and Herdimansyah (1993) |
| Plants | Indonesia | Rubber | Michon and de Foresta (1995) |
| Birds | Indonesia | Rubber, damar, and durian | Thiollay (1995) |
| Birds, bats, butterflies, and dung beetles | Nicaragua | Fallows and pastures | Harvey et al. (2006a,b) |
The belowground ecosystem is out of sight and so often out of mind, but it is very complex and crucially important to the proper functioning of agroecosystems. Lavelle (1996) identified four complex assemblages of organisms with different roles that differ in the size and number of individuals—from the smallest to the largest: the microflora, the micropredators, the litter transformers, and the ecosystem engineers. Agroforestry, as an agricultural system with high aboveground biodiversity and minimum soil disturbance, is recognized to have a diverse rhizosphere associated with the four belowground species assemblages that together form a soil ecosystem typical of a fully functional late successional stage. Tillage, however, disturbs the soil, killing many of the larger organisms and breaking the fungal networks (Cavigelli et al., 2012). This disturbance switches the soil ecosystem back to an early successional stage in which some functions are destroyed and must be rebuilt (Beare et al., 1997).
Forest clearance rapidly changes the soil microflora from one associated with the trees and understory to one associated with the invading pioneers (Mason et al., 1992), with complete clearance of trees having more serious impacts than partial clearance. Complete clearance makes the establishment of tree plantations more difficult even in the moist tropics, as reduced soil inoculum of the appropriate mycorrhizal fungi makes it difficult for planted tree seedlings to develop the best mycorrhizal associations (Mason et al., 1992) and hence survive and thrive. Once established, however, these trees slowly rebuild the soil inoculum over several decades (Wilson et al., 1992).
Importantly, in the few studies that have looked at the effects of conserving biodiversity and the productivity of these systems, there is little, if any, evidence that including biodiversity conservation within these wildlife-friendly perennial crop agroforests has any negative effects on crop production. Indeed, it seems that these agroforests can often successfully combine wildlife and crop production benefits without the need for further deforestation (Clough et al., 2011). In effect, when these complex agroforests also include trees that produce high-value marketable products, replace the natural fallows of shifting agriculture and become commercial fallows, which can be productive and support the livelihoods of local communities for many decades (Michon and de Foresta, 1996), if not a perennial and sustainable sedentary farming system. McNeely & Schroth (2006) recognize that maintaining diversity in agroforestry systems provides a wide range of options for adapting to changing economic, social, and climatic conditions.
So, although this focus on biodiversity conservation within productive farming systems is highly desirable, and indeed the focus of the Ecoagriculture Initiative, it is unfortunate that it mainly focuses on late/mature succession agroecosystems, typically dominated by large trees, and seldom addresses the need to understand the role of biodiversity in the pioneer stages of agroecosystem succession (Leakey, 1999b). One consequence of the predominant focus on mature agroecosystems as habitat for wildlife is that we still have a very poor understanding of the importance of this species diversity in the control of pests, diseases, and weeds. However, recent research has started to fill this knowledge gap by focusing on the agroecological factors that affect the phytopathology of crop productivity, especially in mature agroecosystems.
Regulation processes are complex, but it is recognized that important biological synergisms are a function of plant biodiversity (Altieri and Nicholls, 1999), which, of course, is variable depending on the climate temperature and rainfall, soil types, and levels of disturbance. One important factor in the prevention of disease, pest epidemics, and weed invasions is the limitation of their dispersal. Other factors are the density of herbivores, predators, and other natural enemies; the distance to, and the diversity of, natural vegetation; and the turnover rate of crops within these agroecosystems and the intensity of their management. Furthermore, ecological functions are probably also subject to variation in species abundance, the density dependence of food chains and life cycles, the rates of colonization, reproductive success, mortality rates, and the ability to locate hosts. In addition, there are more species-specific factors, such as odors, repelling chemicals, feeding inhibitors, synchrony of life cycles, etc. One conclusion from agroecological research investigating these factors has been that species composition is more important than the number of colocated species (Altieri and Nicholls, 1999). Thus, identifying the best species assemblages needs to be the focus of future research to determine ecological principles for application under very different circumstances around the world.
Rebuilding Agroecological Functions
In ectomycorrhizal, and perhaps endomycorrhizal, species there are fungi associated with roots of seedling trees, and as the seedling grows, these fungi remain associated with young roots and radiate out away from the tree’s stem, allowing late-stage fungi to colonize closer to the stem. In this way, a successional series of fungi develops over time (Last et al., 1987; Peay et al., 2011). Consequently, for land restoration and stimulation of vegetational colonization, it is important for rapid tree establishment to innoculate tree seedlings with early-stage fungi in the tree nursery prior to planting (Lapeyrie and Högberg, 1994). Likewise, in trees with endomycorrhizal symbionts, the rapid development of these symbiotic relations is important, especially on severely degraded sites where good levels of seedling survival are highly dependent on mycorrhizal inoculation with the best symbionts (Wilson et al., 1991); however, any symbiont seems to be better than none. For agricultural production on severely degraded land, tree establishment is a precursor to the reestablishment of the soil inoculum. This inoculum is then beneficial to the infection and growth of associated crops (Hailemariam et al., 2013). Mulch from leguminous tree species has also been found to encourage the development of soil microflora, as evidenced by increased microbial biomass and increased enzyme activity (Mafongoya et al., 1997; Tian et al., 2001). In Zimbabwe, e.g., actinomycete populations were six to nine times greater when biomass of Vachellia and Calliandra species was applied to the soil surface than when incorporated into the soil (Mafongoya et al., 1997).
Leguminous fertilizer trees have also been shown to promote the return of soil fauna even in highly degraded soils (Sileshi et al., 2008a; Sileshi and Mafongoya, 2006). For example, in Zambia earthworm densities were two to three times higher in maize intercropped with Vachellia, Calliandra, Gliricidia, and Leucaena species compared with maize receiving NPK fertilizer (Sileshi et al., 2008a), whereas maize planted after Sesbania with Tephrosia and pigeon pea fallows had two to three times greater numbers of earthworms than maize alone (Sileshi and Mafongoya, 2006).
Phytopathological knowledge in developing tree-based agroecosystems is substantially based on an understanding of the regulation of pests, pathogens, and weeds in farming systems and on a limited number of case studies (Altieri and Nicholls, 1999; Schroth et al., 2000). In pioneer and early successional stages of agroforestry ecosystems the best-known case studies involve leguminous species. For example, the leguminous shrub Sesbania sesban induces suicide germination of the seeds of the parasitic weed Striga hermonthica, resulting in reduced infestations on cereals (Khan et al., 2007). In other legumes, Desmodium spp. acts as a repellent to stem borers of cereals, whereas Napier grass (Pennisetum purpureum) attracts the pests away from the crops. The combination of these agroecological functions has been described as a push-pull technology (repelling and attracting pests) (Cook et al., 2007). S. sesban has also been credited with reducing the dispersal of maize rust (Krauss, 2004). Some tree and shrub species (e.g., Lantana camara, Melia azedarach, Azadirachta indica, Tephrosia spp.) are known to have insecticidal properties, and some specific insecticidal chemotypes of Tephrosia vogelii have been identified (Belmain et al., 2012). However, there has been concern that some leguminous shrubs also harbor insect herbivores. However, when pure and mixed species legume fallows were tested there was no evidence of serious biomass loss due to herbivory (Girma et al., 2006). Tephrosia fallows had the lowest population densities of 18 species recognized as pests of fallows.
Predator-prey interactions can also be modified in early-stage agroecosystems by diversifying the habitat around fields by planting trees on the field boundaries. For example, in China, pond cypress (Taxodium ascendens) is planted around rice to provide habitat for spiders that control leafhoppers; in Latin America, ants protect cocoa from the arthropod vectors of disease. Trees planted throughout coffee plantations provide habitat for birds, spiders, and ants, all of which contribute to pest control. These properties have been used in farming systems as part of integrated pest management strategies, in which the need for pesticides is reduced by harnessing these natural processes (Dix et al., 1999).
Maintaining Late Successional or Mature Agroecosystems
Recently, a number of studies have started to seek agroecological understanding of pest, disease, and weed problems in mature agroforestry systems in order to find practical, simple, and affordable techniques for poor farmers. These phytopathological studies have been based on both observation and the enumeration of the interactions between certain pests and pathogens and likely predator/parasite species (Table 2.2). Very recently, a few studies have taken observation to the next step in acquiring understanding of specific predator-prey relationships by manipulating parts of the agroecosystem. To some extent, there have been three phases of this research.
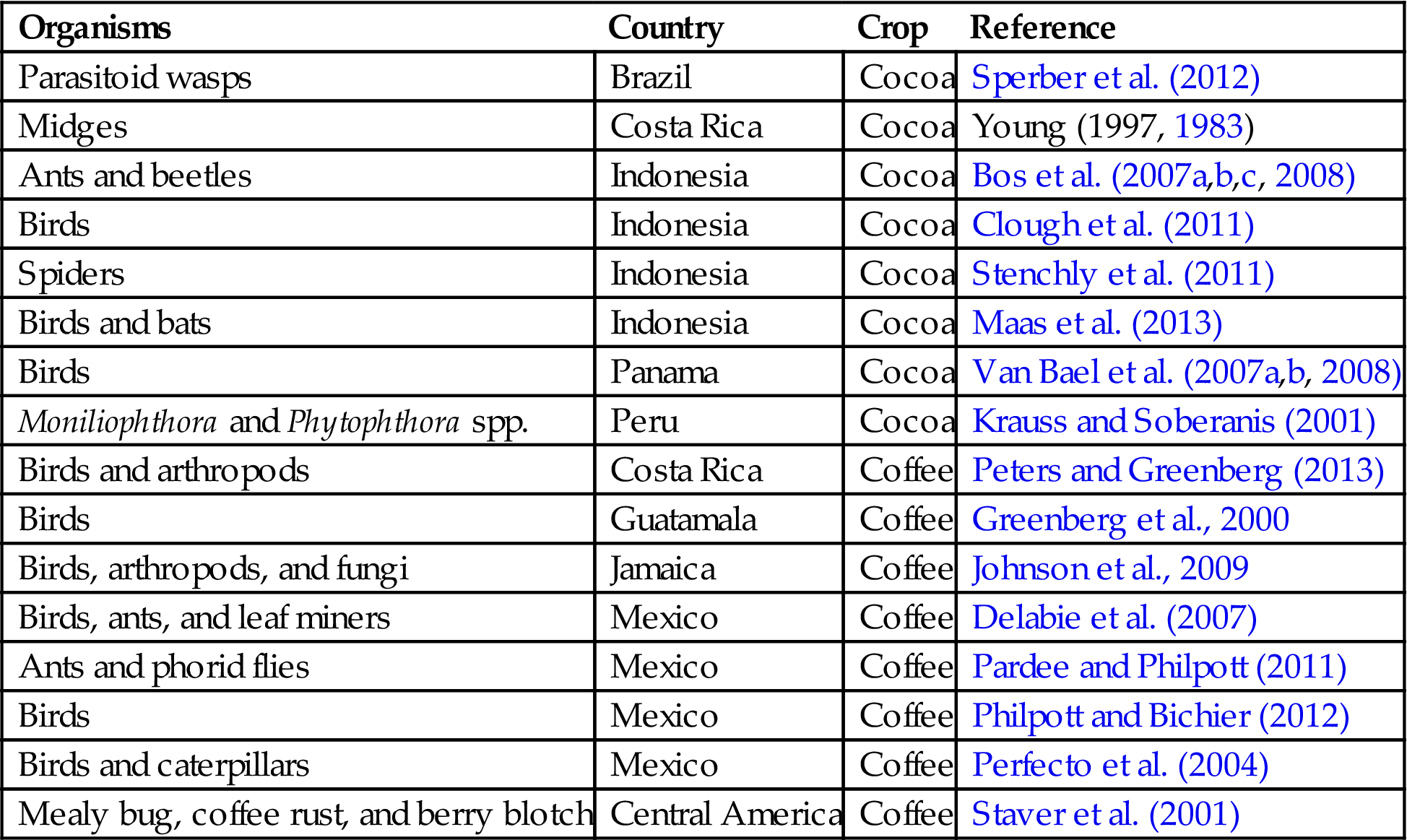
Table 2.2
Published investigations of agroecological functions in agroforestry systems.
| Organisms | Country | Crop | Reference |
| Parasitoid wasps | Brazil | Cocoa | Sperber et al. (2012) |
| Midges | Costa Rica | Cocoa | Young (1997, 1983) |
| Ants and beetles | Indonesia | Cocoa | Bos et al. (2007a,b,c, 2008) |
| Birds | Indonesia | Cocoa | Clough et al. (2011) |
| Spiders | Indonesia | Cocoa | Stenchly et al. (2011) |
| Birds and bats | Indonesia | Cocoa | Maas et al. (2013) |
| Birds | Panama | Cocoa | Van Bael et al. (2007a,b, 2008) |
| Moniliophthora and Phytophthora spp. | Peru | Cocoa | Krauss and Soberanis (2001) |
| Birds and arthropods | Costa Rica | Coffee | Peters and Greenberg (2013) |
| Birds | Guatamala | Coffee | Greenberg et al., 2000 |
| Birds, arthropods, and fungi | Jamaica | Coffee | Johnson et al., 2009 |
| Birds, ants, and leaf miners | Mexico | Coffee | Delabie et al. (2007) |
| Ants and phorid flies | Mexico | Coffee | Pardee and Philpott (2011) |
| Birds | Mexico | Coffee | Philpott and Bichier (2012) |
| Birds and caterpillars | Mexico | Coffee | Perfecto et al. (2004) |
| Mealy bug, coffee rust, and berry blotch | Central America | Coffee | Staver et al. (2001) |
Shade Modification
Tree shade is important to provide the best growing environment for some crops, especially those originating from the forest understory. In cocoa, this shade can be controlled to manage the incidence of diseases, such as frosty pod rot, which is caused by the fungus Moniliophthora roreri (Krauss and Soberanis, 2001). Likewise, in coffee plantations, shade trees can be managed to provide optimal light conditions to minimize the risks from pests [e.g., Cercospora coffeicola (coffee berry and leaf blotch), Planococcus citri (citrus mealy bug), Hemileia vastatrix (coffee rust)] and maximize conditions for beneficial fauna and microflora, even in areas with different soils and climate. Shade has been found to be more beneficial in the dry season and should be reduced by pruning in the wet season (Staver et al., 2001). Predation of insect pests by canopy birds is greatest when the canopy is not intensively managed, with the richness of shade trees explaining much of the variation in bird diversity (van Bael et al., 2007a,b). In another study, disease and insect attacks were more prevalent under single species tree canopies than under mixed canopies, supporting the hypothesis that tree diversity minimizes the risks of pest outbreaks (Bos et al., 2007a,b,c). In addition, shade trees also provide breeding sites for beneficial insects, such as midges, which are pollinators of cocoa (Young, 1982, 1983).
Removal of shade trees has been found to lower the abundance and richness of birds of most guilds, including insectivorous species (Philpott and Bichier, 2012); conversely, the abundance of insectivorous birds was greatest when the canopy cover was dense and species-rich, and there was some dead vegetation. In conclusion, the multifunctional role of shade trees for agriculture and biodiversity conservation is now recognized, but their important role in risk avoidance from insect pest outbreaks is inadequately understood. Nevertheless, it is clear that a diversified food-and-cash-crop livelihood strategy is possible (Tscharntke et al., 2011).
Bird Exclusion
A small number of experiments have investigated the importance of bird and bat populations on organisms further down the food chain in multistrata cocoa and coffee agroforestry systems by setting up exclusion experiments (Johnson et al., 2009; Maas et al., 2013; Peters and Greenberg, 2013). An experiment in which caterpillars were placed on coffee plants with and without bird exclusion confirmed that birds do reduce pest outbreaks, but this was only significant in association with high floristic diversity, indicating the importance of species diversity in the habitat (Perfecto et al., 2004). Likewise, Greenberg et al. (2000) found that exclusion of birds also resulted in a 64–80% reduction in large (>5 mm) arthropods and consequently some increased herbivory. Other studies in cocoa and coffee have generally confirmed these findings, showing that exclusion does generally increase the abundance of foliage-dwelling insect herbivores, with knock-on impacts on leaf damage (van Bael et al., 2007a,b, 2008). In Indonesia, the exclusion of birds and bats resulted in a 31% reduction of crop yield, illustrating the economic importance of these insectivores (Maas et al., 2013).
Food Chain and Life Cycle Studies
Shaded coffee and cocoa agroecosystems with a well-developed canopy of shade trees have high biodiversity, a wide diversity of microclimates and ecological niches (Siebert, 2002), and, as we have seen, fewer pest problems, most likely because of the greater abundance and diversity of the predators of herbivores. However, the species interactions in predator or parasite impacts on prey and crop production can be very complex. For example, without shade phorid flies negatively affect ant populations, and because ants negatively affect coffee berry borer, there is more damage to coffee berries (Pardee and Philpott, 2011). This indicates how shade impacting on phorid flies can enhance the biological control of berry borer by ants. Another example of the shade benefits involves parasitic wasps of the coffee leaf miner. Leaf damage by the miner was lower when twig-nesting ants were abundant, indicating that ant populations control leaf miners in shaded coffee (de la Mora et al., 2008); thus, through the effects of shade, ants provide important ecosystem services in coffee agroecosystems. Perhaps similarly, a study of parasitoid wasps in cocoa agroforests has found that wasp community composition was influenced by season, disturbance, the species richness of the shade trees, and the scale of the cocoa planting (Sperber et al., 2012). However, the impacts of such interactions can be complex and affected by such things as the climate zone and feeding habits of the bird species. For example, large frugivorous and insectivorous birds are more attracted to agroforests than small-to-medium insectivores and nonspecialist feeders. As might be expected, frugivores and nectarivores favored canopies with high species richness (Clough et al., 2009a,b). Thus, studies of bird populations need to take into account species from different functional groups (Sekercioglu, 2012).
Other studies in cocoa agroforests have found that canopy thinning leads to a reduction in the diversity of forest ant species but not of beetles (Bos et al., 2007a,b,c). This contrasting effect of canopy management illustrates the different effects of shade on different organisms and the need for a better understanding of food chains and life cycles in agroecosystem functions. For example, web-building spiders may be important predators. A study by Stenchly et al. (2011) found that web density in cocoa agroforest was positively related to canopy openness and that line and orb webs were the most common type. The presence of Philidris ants was also positively associated with orb-web density but not with other web types. The practical importance of these studies is that fruit losses due to pathogens and insect attacks have been found to be greater when the tree diversity of the canopy is low. This supports the hypothesis that diversity in the upper strata of agroforestry systems has beneficial effects on agroecological functions (Bos et al., 2007a,b,c).
Meta-analysis of the biodiversity and ecosystem service benefits of cacao and coffee agroforests has confirmed the wide-scale benefits of shade trees for the provision of habitat for pest control by insectivores in Latin America and Asia, and less-well-studied Africa (de Beenhouwer et al., 2013; Poch and Simonetti, 2013). However, it is quite possible that these studies have only started to explore the ecological interactions of importance in sustainable agriculture and so further ecological research is needed.
To gain a better understanding of the complex life cycle and food chain interactions between species in mixed species plantings, such as agroforests, recent studies have established some long-term experiments involving species mixtures that vary in the diversity of the planned biodiversity (the planted crops and trees), their species densities, and their configurations. This creates very different microclimates (irradiance, temperature, humidity, etc.) within both the ground flora and the canopies, which in turn affects the colonization of the ecological niches so formed (Siebert, 2002). To try to develop some deeper understanding of the importance of all this variability on the incidence, severity, and dynamics of pest, disease, and weed organisms, Leakey (2012b) has proposed two experimental designs. These experiments were designed to explore agroecosystem breakdown with regard to witches’ broom disease in cocoa.
These designs seek to answer some multidisciplinary research questions spanning the agroecological successional from pioneer to mature phases, i.e., from planting through to maturity (Table 2.3), including the impacts of the agroecosystem on the health, yield, and sustainability of cocoa production. It is recognized that in the long-term the results of experiments like these will probably be very different depending on the level of land degradation at the planting site and its distance to mature forest. It is also recognized that such experiments are extremely difficult to implement without the confounding effects of competing root systems between juxtaposed treatments; thus individual replicates of each treatment are surrounded by guard rows of similar species composition.
Table 2.3
Research questions aimed at an examination of the interactions between planned and unplanned biodiversity in cocoa agroforests.
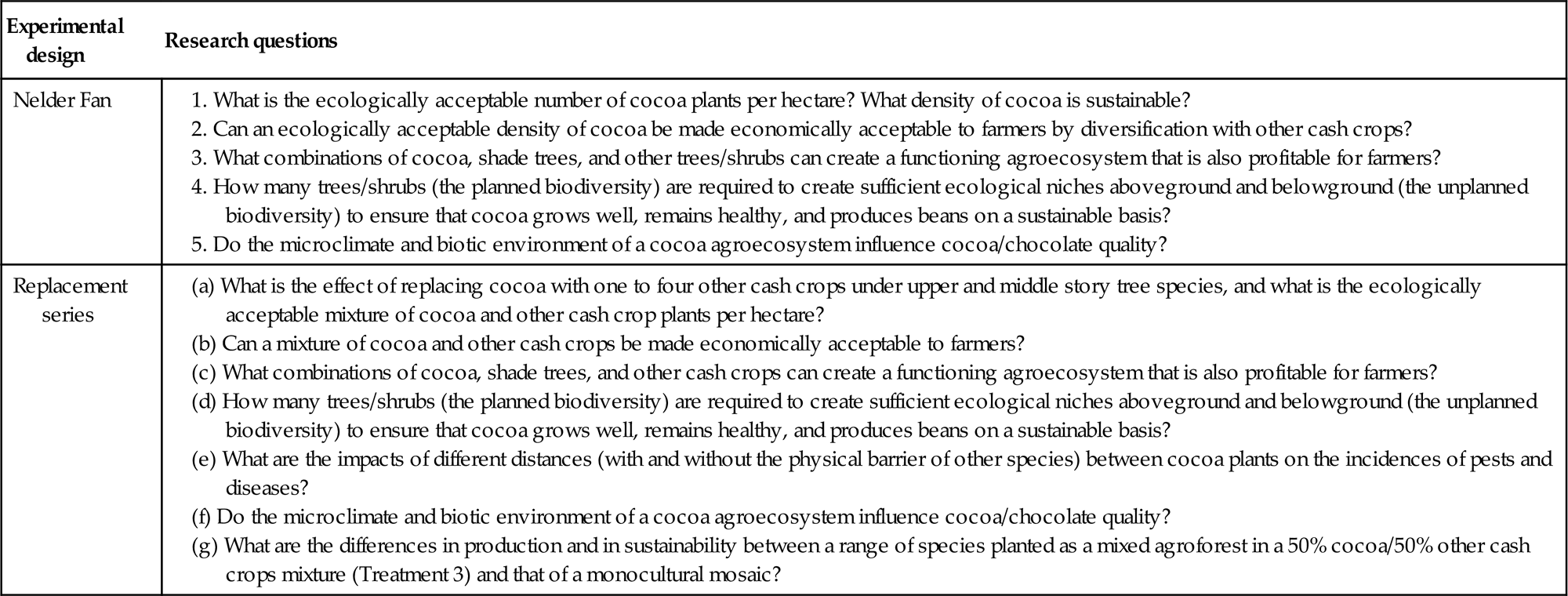
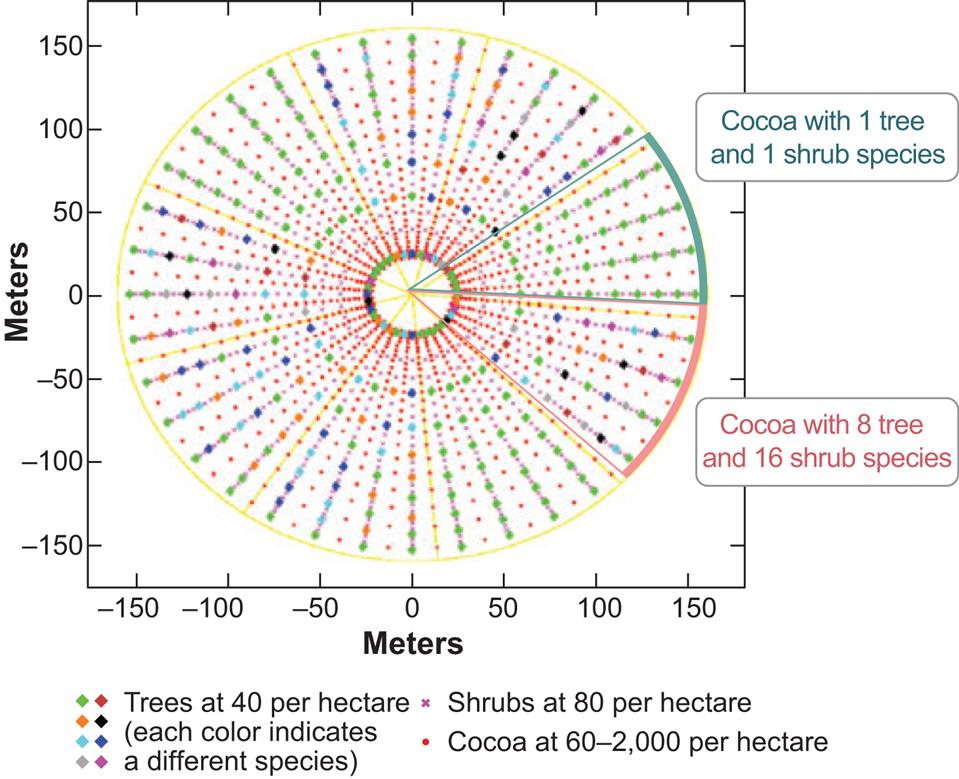
The first design involves a complex multistrata agroforestry experiment laid out as a Nelder Fan (Nelder, 1962), and the second experiment is a complex replacement series (de Wit, 1960). Specifically, these designs seek to examine the relationships in mixed species cocoa agroforests between spacing, microclimate, the planned and unplanned biological diversity, the incidence of pests and diseases in cocoa, and the overall production and economic returns from cocoa and the other companion trees. The replacement series study adds an assessment of the impacts of different configurations of companion species grown in differing proportions within the canopy. Both these experiments are also examining how a range of indigenous companion crops producing agroforestry tree products can additionally provide income generation and livelihood benefits.
The Nelder Fan design aims to examine some ecological questions (Table 2.3) that arise from growing a single crop (cocoa) at different densities (60–2000 trees per hectare) under a constant density of a canopy composed of either one understory shrub species and one canopy tree species or of 16 understory shrub species and eight canopy tree species (Fig. 2.1).
The second design modifies the replacement series design to test the replacement of cocoa with five other cash crops under a canopy of five tree species and five subcanopy species (Fig. 2.2). The upper-story trees and subcanopy trees/shrubs are at the same spacing (9 m×9 m), each of which is surrounded by eight cocoa plants, two plants each of four cash crops, or a mixture of four cash crops with four cocoa (3 m×3 m), depending on the treatment within the replacement series. The modular structure of the canopy ensures that the physical properties of the multistrata agroforest (i.e., the three stories of the canopy) remain constant across the experiment. To act as a control treatment in the replacement series, each of the different species is grown as a monoculture and at the same spacing as it occurs in the 50:50 treatment of the replacement series. This allows the comparison of production and income per unit area from a monoculture with that of the integrated species mixture.
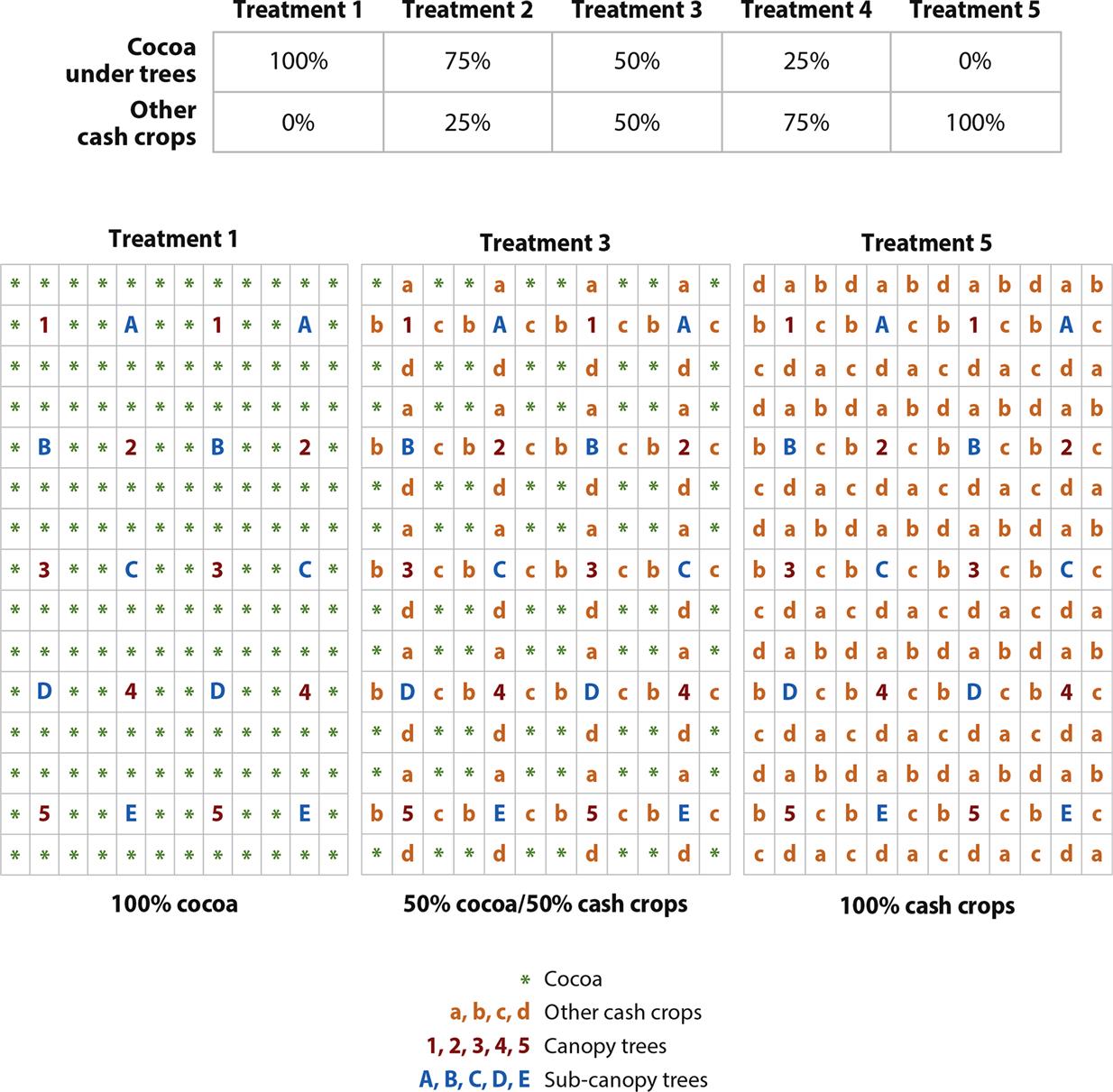
In both of these experiments, the design embraces the idea that the canopy and subcanopy trees should provide marketable products in addition to shade and other environmental/ecological services. This is in recognition that in addition to minimizing the risks of pest, disease, and weed problems in commodity or food crops, farmers want to increase their income and improve their livelihoods. For the past 20 years, the domestication of such trees has been aimed at this poverty-alleviating benefit (Leakey et al., 2012).
Filling the Niches Below the Canopy With Useful Plants
Agroforestry often creates opportunities for shade-adapted species to fill shady niches and increase the benefits derived from mixed cropping systems. In this connection, most existing food crops have been selected and bred for cultivation in full sun, so there are opportunities for plant breeders and domesticators to develop new crops or crop varieties that are better adapted to partial shade (Leakey, 2012b), e.g., Eru (Gnetum africanum) in Cameroon. In other initiatives, socially and commercially important herbaceous species are also being domesticated as new crops to meet the needs of local people for traditional foods and medicines as well as other products, and these too can fill niches in complex species mixtures (Okafor, 1980; Schippers, 2000; Smartt and Haq, 1997). In addition to these ecological benefits, this diversification also has social and economic benefits that go a long way toward meeting the goals of multifunctional agriculture.
Landscape and Scaling Issues
With little new land for the expansion of agriculture without the loss of now scarce forest, it is clear that the agroecology of agroforestry systems is an important area for more research (Siebert, 2002). This need is probably further expanded by the complexity added by scaling up to the level needed for ecological equilibrium in agricultural landscapes. In practice, the achievement of scale may need to depend on the formation of land-use mosaics with corridors between forest patches (Leakey, 1999b). This is illustrated by the finding that the diversity of frugivorous and nectarivorous birds decreased with increasing distance from the forest, whereas granivorous birds increased in diversity with increasing distance from the forest (Clough et al., 2009a,b). This illustrates that species of different functional groups differ in their need for a forest resource in the landscape. This is of importance both for wildlife conservation and almost certainly for the optimization of agroecological interactions.
In Central America, coffee agroforests are important habitat for both sedentary and migratory birds of regional and international importance. However, the role of agroforests on migratory bird populations and of migratory birds on agroecological functions are not well understood. However, one study has found that the effects of bird predation were especially strong when bird diversity was high as a result of the presence of migratory birds. Importantly, however, this period of high bird diversity was associated with greater reductions of arthropod density (van Bael et al., 2008).
Fortunately, when considering landscape issues there is now increasing information about the tree species richness and composition of smallholder farming systems around the world (Goenster et al., 2011; Kindt et al., 2006a,b; Michon and de Foresta, 1996) and what it means for production, farm management, and farmer livelihood, and vice versa (Carsan et al., 2013). Of course, the farmer is much more interested in the livelihood benefits from agroforestry systems than in the wildlife per se. Thus, when engaging farmers in discussions about agroforestry, it is important to explain the livelihood benefits from complex agroforests and the need to utilize different tree species that produce useful and marketable products as the structure of the mature agroecosystem, i.e., the planned biodiversity (Perfecto and Vanderdeer, 2008). As we have seen, these perennial crops then create niches aboveground and belowground for the wild organisms—unplanned biodiversity—which are vital for the completion of complex food chains and the closure of numerous interactive life cycles at different trophic levels. These functions, together with nutrient, carbon, and water cycling, are the basic processes determining the functioning of ecosystems.
The Big Picture: The Role of Agroecology and Agroforestry in Tropical Agriculture
As mentioned in the “Introduction” section, deforestation and land degradation are now seriously affecting both agricultural crop yields in the tropics and the availability of productive land. This, together with the abject poverty of many smallholders unable to purchase artificial fertilizers and pesticides, makes the importance of land rehabilitation by agroecosystem restoration a prime objective for rural development (Leakey, 2012b). How can this be done? Two literature reviews concluded that it is possible for productive agriculture to provide both food security and biodiversity using appropriate agricultural practices that support functioning agroecosystems (Chapell and LaValle, 2011; Tscharntke et al., 2012).
One suggestion for how to simultaneously resolve the issues driving food and nutritional insecurity, poverty, and environmental degradation is to take a three-step agroforestry approach (Fig. 2.3). This combines biological nitrogen fixation with the diversification of the farming system for the restoration of agroecological functions and then adds to that the generation of income from the domestication of indigenous tree species that produce useful and marketable products for local and regional trade (Leakey, 2010, 2012e, 2013). These three steps are also aimed at closing the yield gap (the difference between potential yield of a food crop and the actual yield achieved by farmers) in staple food production by increasing soil fertility and generating income for farmers.
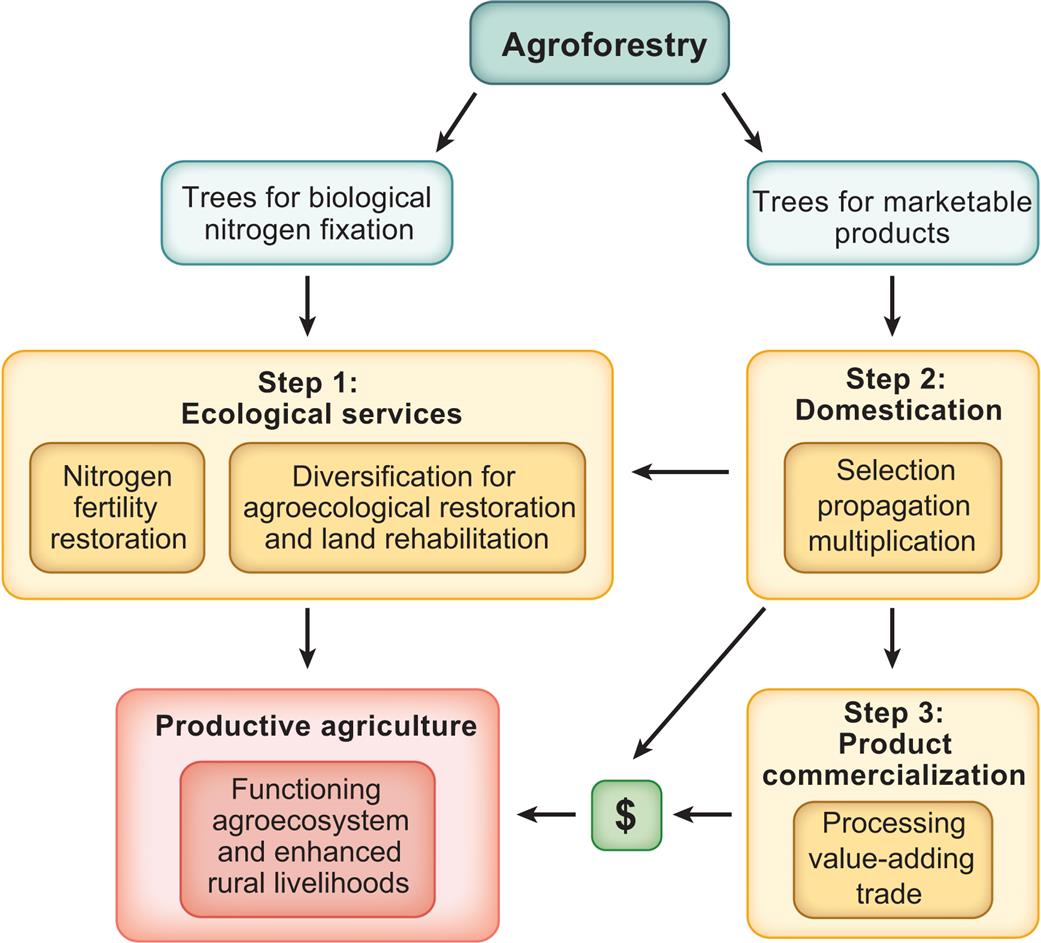
Ecological Restoration (Step 1) builds on over 25 years of research experience, led by the ICRAF on the use of a number of leguminous nitrogen-fixing trees and shrubs, in improved fallows, relay cropping, and, more recently, evergreen agriculture. As mentioned earlier in the section on addressing soil fertility, field experience indicates that twofold to threefold increases in crop yield are typical after 2 years of these nitrogen-fixing tree fallows. Even on a small farm, this yield increase is enough to free up space to diversify the cropping system with other income-generating crops or farm enterprises and initiate agroecological restoration. Then adding trees which produce marketable products further diversifies the farming system and forms a productive tree canopy that provides shade to cocoa and coffee. This leaves room for other crops in the canopy gaps and in the landscape mosaic.
Tree domestication (Step 2) by selecting and vegetatively propagating their elite trees for planting as superior cultivars to produce culturally and traditionally important and nutritious foods (Asaah et al., 2011; Leakey and Asaah 2013; Tchoundjeu et al., 2006, 2010), has, over the past 20 years, made great progress in assisting poor rural communities.
Product commercialization (Step 3) creates business and employment opportunities in cottage industries engaged in processing, value adding, and marketing of the products of these agroforestry trees.
By combining agroecological restoration with income generation within a participatory integrated rural development program that provides community training and education in a wide range of relevant skills (Degrande et al., 2012), agroforestry becomes a powerful new tool to address the cycle of land degradation and social deprivation. The income-generating component adds critical value to agroecology and may even be an important incentive for farmers to diversify their farming systems. Consequently, a collective action initiative in Cameroon (Gyau et al., 2012) is now being seen as an approach to delivering multifunctional agriculture and sustainable intensification to tackle the big issues of poverty, malnutrition, and hunger. If implemented on a sufficient scale, this approach also has potential to mitigate climate change because the trees, and their soils, sequester substantial quantities of carbon. These lessons for integrated rural development have now been drawn together as a set of 12 Principles and Premises (Leakey, 2014f) based on the agroecological role of trees for upscaling and wider implementation of the sustainable intensification of agriculture.