
Agroforestry and the Mitigation of Land Degradation in the Humid and Sub-Humid Tropics of Africa
This chapter was previously published in Cooper, P.J.M., Leakey, R.R.B., Rao, M.R., Reynolds, L., 1996. Experimental Agriculture, 32, 235–290, with permission of Cambridge University Press
Abstract
Population pressures, exacerbated by a range of social and political factors, have resulted in widespread deforestation, land degradation and the cultivation of marginal and sloping land. The resultant declining soil fertility and soil erosion threaten the sustainability of small-scale farming, especially as external nutrient inputs are seldom available to farmers – with additional shortages of tree products. This review focuses on research to enhance soil fertility, prevent soil erosion, provide high-quality dry season fodder and much-needed income from marketable products. The results are encouraging, both in technical performance and farmer enthusiasm. However, to ensure that agroforestry solutions are locally relevant, it is essential that farmers are included as research partners. We are optimistic that agroforestry provides relevant and useful solutions for today’s land degradation problems and that these will evolve in both diversity and intensity to effectively meet the needs of Africa.
Keywords
Agroforestry; domestication; erosion; land degradation; rehabilitation; soil fertility; smallholder farmers; sustainable agriculture; tree products
Summary
In the last 35 years, the population of sub-Saharan Africa has increased nearly threefold and is expected to reach 681 million by the year 2000, with nearly 50% of the population living in urban centers. Such population pressures, exacerbated by a range of social and political factors, have already resulted in widespread land degradation in areas of high population densities and the expansion of agriculture on to marginal and sloping land. Declining soil fertility and soil erosion are increasingly threatening the sustainability of small-scale farming systems throughout Africa, and affordable external nutrient inputs are seldom available to farmers. In addition, shortages of wood for construction and fuel and high-quality dry season fodder for livestock are widespread and serious constraints to farm productivity.
Agroforestry, the deliberate integration of woody perennials into crop and livestock systems, has the potential to mitigate many of these constraints through both the service and production functions played by trees. In recent decades much agroforestry research has been undertaken in sub-Saharan Africa. In this review we focus specifically on research that addresses the potential of agroforestry systems to enhance soil fertility, prevent soil erosion, provide high-quality dry season fodder or generate much-needed income through the production of high-value goods.
Much emphasis has been placed on a wide range of agroforestry systems for the maintenance of soil fertility and the prevention of soil erosion losses, and encouraging results, both in technical performance and farmer enthusiasm, have occurred. However, it is clear that agroforestry solutions to land degradation are always likely to be location-specific in their relevance, performance, and farmer acceptability. It is essential that farmers are included as research partners to determine what is appropriate for their conditions.
Good progress has also been made on identifying fast-growing leguminous trees and shrubs for high-quality livestock fodder supplements. Where livestock enterprises, such as peri-urban milk production, are market-oriented, the adoption and impact of such systems have been high. Given population and urbanization projections, it is likely that fodder trees and shrubs will have a major role to play in meeting future feed demands for both milk and meat production. Research on the potential of high-value indigenous and exotic trees to generate income has been less extensive in Africa, although the huge potential of this approach has been clearly demonstrated by farmers in Southeast Asia. We suggest that there is a need for increased research emphasis on the domestication of high-value indigenous trees and their integration into more sustainable, diverse and intensive land-use systems.
We conclude that, although good progress has been made in agroforestry research in Africa and farmer adoption is occurring, future population projections pose a clear challenge. Agroforestry systems that provide solutions for today’s land degradation problems will need to evolve in both diversity and intensity if they are to remain relevant and effective for tomorrow’s Africa.
Land Degradation
It is estimated that the population of sub-Saharan Africa has nearly trebled in the last three and a half decades, rising from 210 million in 1960 to 495 million in 1990, and it is predicted that it will reach 681 million by the year 2000 (UNFPA, 1992). In long-term predictions of population growth, Bulatao et al. (1990) estimate that the population of Africa as a whole will continue to increase well into the 21st century, reaching a maximum of around 3000 million by the year 2100. It is true that these figures are based on estimates and projections that exclude the impact of natural disasters, wars, and disease, and hence must be treated with caution. Furthermore, they are not specific to the humid and subhumid tropics of Africa. However, they do clearly illustrate the alarming situation of unprecedented population increases that are likely to occur, and which are already occurring, particularly in the higher-potential humid and subhumid regions.
The impact of the already huge increases in population on the degradation of the natural resource base has been as predictable as it has been devastating. This devastation has arisen from a complex interaction of biophysical, socioeconomic, and political factors. For example, as population densities have increased, traditional and sustainable systems of shifting cultivation have suffered from shorter and shorter fallow periods of natural forest or woodland regeneration (Fig. 4.1). This situation has eventually given way to sedentary agriculture on small-scale land holdings. In addition, continuing population pressure has resulted in further forest clearance for agricultural land. From a study of 40 countries in Africa, Dembner (1991) estimates that 50 million hectares of land was deforested between 1980 and 1990. The possible future impact of 3000 million people in 2100, if steps are not taken to reduce the rate of deforestation and land depletion, is hard to imagine.

High rates of urbanization in Africa have also exacerbated land degradation. The growing numbers of urban dwellers require food, fuelwood, and timber products. These needs result in a steady transport from agricultural land to urban settlements of nutrients that end up as useless and unrecycled waste. In Africa as a whole, the proportion of people living in urban settlements has risen from around 15% in 1950 to an estimated 40% in 1995 and is expected to reach 54% by 2025 (UNEP, 1992). Nutrient export from agricultural land to urban centers is, however, not the only source of soil-nutrient depletion, particularly with regard to nitrogen (N). In analyses of nutrient budgets of cropped fields on small-scale mixed farms in Western Kenya, Shepherd et al. (1994) estimated that in such high-rainfall areas (1600–1800 mm a−1), 63 kg N ha−1a−1 were lost through leaching, denitrification, and volatilization. This can be compared with 43 kg N ha−1a−1 lost through export from the field in crop yield. For the range of contrasting farm management options they examined, net nitrogen balances for cropped land ranged from −39 to −110 kg ha−1a−1 and net phosphorus (P) balances from −7 to +31 kg ha−1a−1. For the sub-Saharan region as a whole, it has been estimated that the annual average loss per hectare is 22 kg N, 2.5 kg P, and 15 kg K (Stoorvogel et al., 1993).
A further result of population increase and the nutrient depletion of potentially productive land has been the expansion of agriculture on to marginal sloping land and into drier areas traditionally reserved for livestock grazing. Such lands not only suffer from nutrient export but unless carefully managed are especially prone to wind and water erosion of the topsoil where the bulk of soil nutrients available to crops is located. It is estimated that steeply sloping land accounts for about 10% of the geographical area of sub-Saharan Africa, although in some countries in east and southern Africa, such lands constitute as much as 40–70% (Fig. 4.2). Human-induced water erosion accounts for nearly 50% of land degradation in Africa (WRI, 1992).

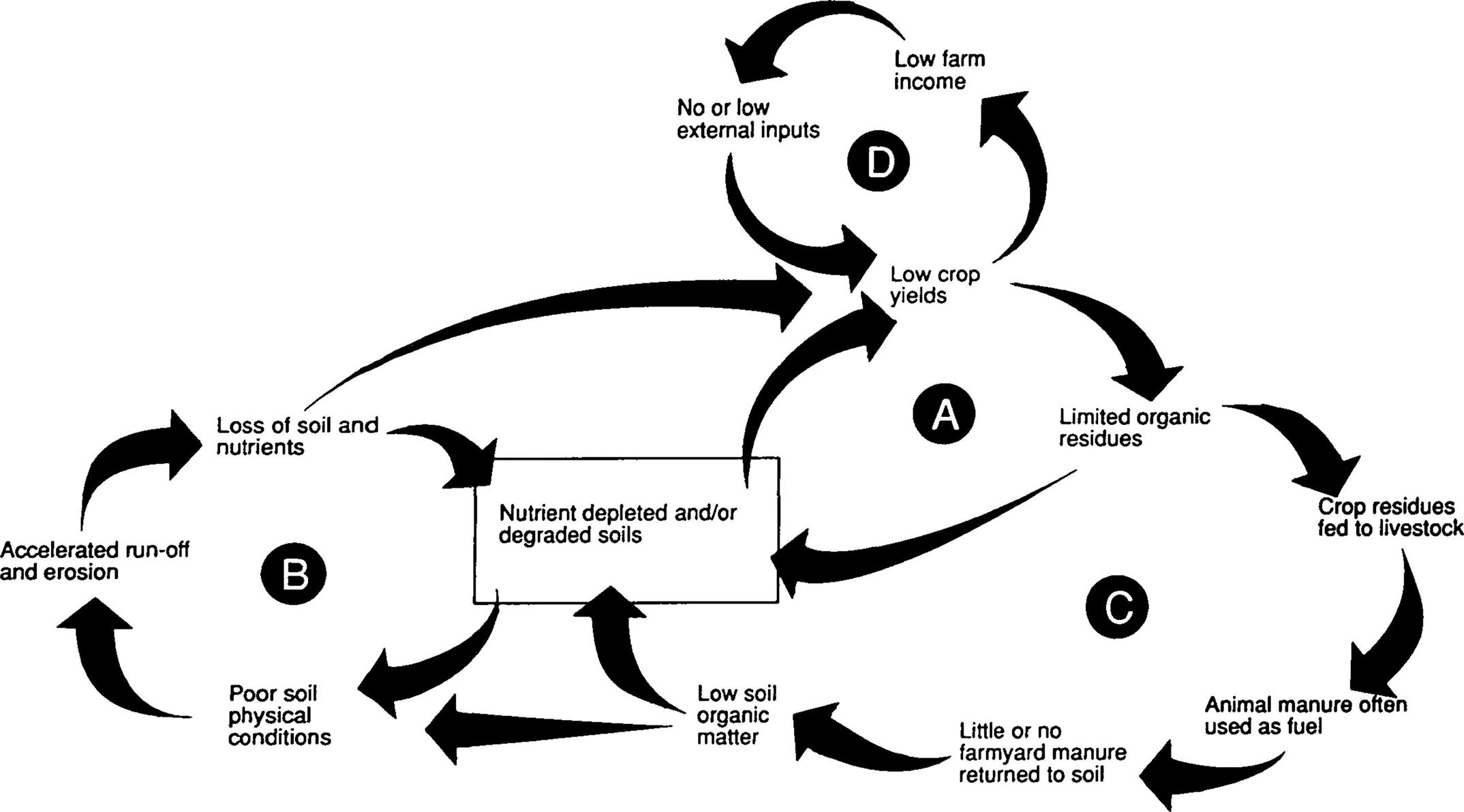
Few small-scale farmers can afford to replenish this steady loss of soil nutrients with external inputs of organic or inorganic fertilizers. Often fertilizer is simply not available or too expensive and in many instances farmers are forced to use animal manure as fuel rather than as fertilizer. They cannot afford to implement long-term soil conservation and improvement strategies that do not provide short- or medium-term benefits. Many of these problems have been exacerbated by, e.g., insecure land tenure, land fragmentation, lack of access to credit, pricing policies that favor urban consumers, inappropriate trading policies, and poor access to both local and international markets. These problems have all fueled the downward spiral of land degradation (Fig. 4.3). The results of these combined forces have been the development of a range of major production constraints now facing small-scale farmers.

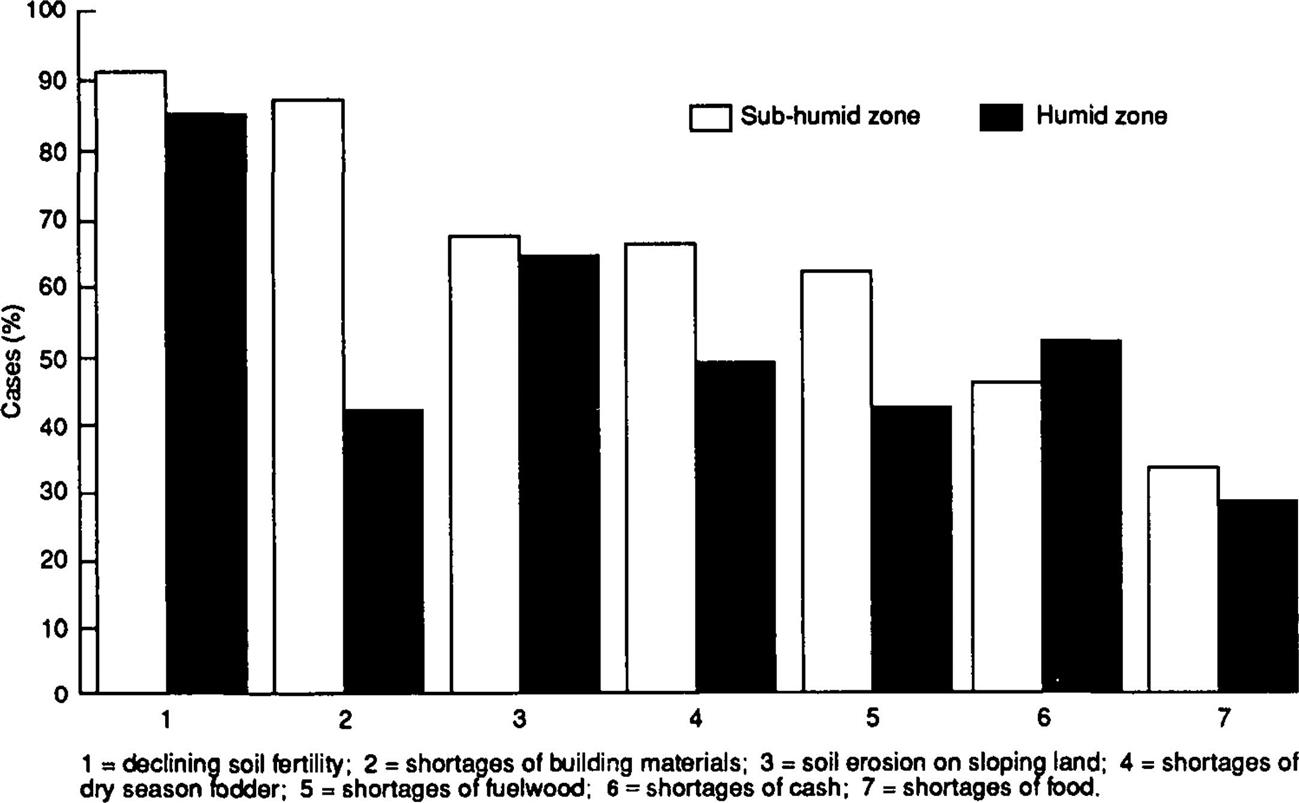
Raintree (1987) examined the relative importance of these constraints by summarizing the output of ICRAF’s (the International Center for Research in Agroforestry, now called the World Agroforestry Center) diagnosis and design (D&D) exercises (equivalent to participatory rapid rural appraisal) undertaken in 47 land-use systems in 9 countries of sub-Saharan Africa. These D&D exercises asked farmers to list the major constraints they faced (Fig. 4.4). What is immediately apparent is that farmers were aware of both declining soil fertility and soil erosion as important constraints to their welfare. Further, it is interesting that shortages of building materials and fuelwood, a direct consequence of forest and woodland clearing and the denudation of the landscape of trees, appear to be of more concern in the subhumid than in the humid regions. This is associated with both the slower regenerative capacity of woody vegetation in the drier areas and greater population densities. Fodder shortages are also an important constraint, particularly in the subhumid regions. This reflects the greater livestock numbers, the more severe dry season and the continuing destruction of natural grazing lands for agriculture. Given declining soil fertility and hence declining crop yields, it is perhaps surprising that farmers did not mention food shortages and cash shortages more frequently, but this probably reflects the subsistence nature of many small-scale farms in sub-Saharan Africa and their ability to find other sources of family income through off-farm employment.
In conclusion, if we accept the postulated population growth estimates, the prospects for future generations of the urban and rural people of Africa are indeed bleak. What can be done to alleviate this crisis? This chapter attempts to draw together evidence from biophysical, social, and environmental studies to illustrate that agroforestry, if applied on a large enough scale, does have the potential to help alleviate the problems. It is, however, self-evident that, unless backed by supportive national and international policies and social motivation, farmers will continue to struggle to make a living in ways that lead to land degradation, rather than adopting improved land management techniques.
The Role of Agroforestry
In order to place agroforestry in a context with other land-use systems, ICRAF has adopted the following definition:
Agroforestry is a collective name for land-use systems and practices in which woody perennials are deliberately integrated with crops and/or animals on the same land management unit. The integration can be either in a spatial mixture or in a temporal sequence. There are normally both ecological and economic interactions between the woody and nonwoody components in agroforestry.
Raintree (1987) noted that a narrow interpretation of this definition would require the actual physical interaction of trees with herbaceous crops, pastures, or livestock in space or time. However, from a problem-solving point of view a broader and more useful interpretation allows trees to be grown on land management units ranging from a field, to a farm, a community, or a watershed. This more liberal interpretation, which is increasingly being adopted, only requires that there be interactions between the components. For example, at the farm level, a block planting of a woodlot, fodder bank, or a discrete home garden can each be considered as an agroforestry system. This is by virtue of their economic interaction with other components of the farm in drawing on the same pool of family labor, capital and management resources.
At the farm level, trees have the potential service functions of soil fertility improvement and/or maintenance, the reduction of wind and water erosion, enhanced microclimate, livestock containment through living fences, and demarcation of internal and external boundaries. On a larger-scale, trees can improve the hydrological cycles in watersheds, and play an important role in the maintenance of soil, insect, plant, and wildlife biodiversity. Trees in agroforestry systems also have a potential role in the sequestration of carbon in above- and belowground biomass and enhancing levels of soil organic carbon. Unruh et al. (1993) estimated that 1550×106 ha of land are suitable for some kind of agroforestry intervention in sub-Saharan Africa. If this potential were fully realized, they calculated that between 8 and 54×1015 g of carbon would be sequestered, enough to offset global emissions of carbon from fossil fuels for only 1.7–9 years. They concluded that the secondary effects of agroforestry in reducing rates of deforestation and hence emission of carbon may be more important than the primary effects of carbon sequestration.
From a production perspective, trees provide fuelwood, building materials, and high-quality dry season fodder. They are also an important source of income generation through the provision of high-value products such as poles, timber, fruits, medicines, resins, and gums. There is increasing awareness of farmers’ indigenous knowledge and uses of trees. For example, ICRAF (1992a) recently presented detailed information of 32 distinct service and production functions of over 170 indigenous and exotic species found in Kenya alone. Among scientists, there is also greater awareness of the potential to domesticate these species (Leakey and Newton, 1994a,b). Agroforestry, through both the service and production functions of trees, therefore has the potential to alleviate or, in some instances, remove completely many of the principal constraints recognized by farmers which result directly from land degradation.
In this review, we shall deliberately focus on the role of trees in mitigating land degradation at the farm and field levels. It is at this level that the bulk of current information is available, and it is farmers who are and will be the prime instigators of improved land management. Farms are the most important building blocks from which larger-scale impacts will be achieved. As previously indicated, there are many interlinked and interactive causes and processes involved in the downward spiral of land degradation (Fig. 4.5). We shall structure our review accordingly and provide evidence on the capacity of agroforestry to break this downward spiral through soil fertility improvement (intervention point A), the reduction of soil erosion (point B), the provision of dry season fodder (point C), and generating income (point D).
Soil Fertility Improvement
It has long been recognized that trees have the potential to increase nutrient availability to crops, a fact which is self-evident given the historical sustainability of shifting cultivation throughout the humid and subhumid tropics. Some forms of agroforestry attempt to mimic the role of secondary forest or woodland fallows through the deliberate inclusion of fast-growing leguminous trees into the farming system for the maintenance or improvement of soil fertility. Various systems are being evaluated which include trees grown either in spatial mixtures or in temporal sequences with crops. Several processes have been identified by which trees can enhance the chemical and physical properties of soils (Ingram, 1990). They include:
1. Nitrogen input into the system through biological nitrogen fixation by rhizobial bacteria in root nodules of certain tree species.
2. Deep nutrient uptake by trees from below the crop rooting zone and subsequent surface deposition as litter, biomass burning, or prunings. In addition, belowground litter of decaying root systems may be important.
3. A more closed nutrient cycle through the capture of nutrients which would otherwise be leached from beyond the tree/crop rooting zone.
4. Improved soil physical conditions (e.g., better aggregation, reduced bulk density, improved infiltration) resulting from higher levels of organic matter, old tree root channels, and increased macrofaunal activity.
5. Reduced aluminum (Al) toxicity and low pH through enhanced cycling of bases and the production of metabolic compounds that temporarily complex Al.
6. Increased availability of P through mycorrhizal associations.
7. Improved activity of soil organisms (such as fungi, arthropods, termites, and worms) through a cooler and moister microclimate.
Early research in agroforestry largely concentrated on evaluating “systems” for improving soil fertility with less emphasis on trying to assess the relative importance of the possible biophysical processes involved. Such research has in many instances been invaluable in defining “what works where” but has been less successful in explaining “why.” Given the considerable complexity of above- and belowground interactions, and the technological challenge of measuring discrete processes in agroforestry systems, there is still some way to go. Recent reviews of process-oriented agroforestry research have identified some hard data (Ingram, 1990; Anderson and Sinclair, 1993) but Sanchez (1996), in discussing the evolution of agroforestry as a science, concluded: “Agroforestry is not there yet. Its underlying principles are yet to be developed in a sufficiently rigorous manner that assures predictive understanding.”
Given the ongoing development of these principles, we will concentrate on presenting examples where rigorous “systems-oriented” research, both on-station and in farmers’ fields, has made a start in demonstrating what works, or does not, and where.
Hedgerow Intercropping
Hedgerow intercropping (synonymous with alley cropping) is perhaps the most widely researched agroforestry system in sub-Saharan Africa (Kang et al., 1990). Trees are grown as spatial mixtures in hedges, typically spaced 4–6 m apart with a within-hedge spacing of 0.25–0.5 m. Crops are grown between the hedges and tree biomass, obtained through periodic pruning during the cropping period, is added to the soil as green manure. Early work on this system was pioneered on Alfisols and Entisols by the International Institute for Tropical Agriculture (IITA), and appeared very promising (Kang et al., 1981). Since that time scientists throughout sub-Saharan Africa have evaluated this system under a wide range of soil and climatic conditions. Results have been mixed and in general disappointing. Recent reviews of hedgerow intercropping trials reported in the literature (e.g., Akyeampong et al., 1995) have highlighted some of the following important factors.
First, many of the earlier trials suffered from experimental design faults, which could have resulted in an upward bias of the yields reported in hedgerow intercropped plots and reduced yields in the control plots. These have been summarized by Coe (1994), who identified 10 problems commonly encountered in research reported by ICRAF as well as other institutes. Principal among these were:
1. Experimental plot size too small, resulting in above- and belowground interference between treatments. For example, one experiment in Kenya which compared three tree species had tree roots of all three species in every plot after 4 years. Such an effect is perhaps even more important when tree roots invade adjacent “no tree” controls. The trees get extra soil resources from an area perhaps twice that of the hedgerow plot, and these soil nutrients are then concentrated as green manure within the hedgerow treatment area. Not only does this result in unrealistically high amounts of nutrients added as green manure but also control plots are depleted of nutrients and water. Coe quotes an example where nearby yields from sole maize plots outside the hedgerow intercropping experimental area were two to three times higher than the sole maize control within the experiment, even though the management of both plots was identical. More recent experiments have to a large extent overcome these problems through the use of much larger plots and the installation of root barriers or root trenches around control plots. However, such barriers must be reinstalled frequently since tree roots have been shown to grow under them and up into the control plots within 1 year (Singh et al., 1989).
2. Many experiments have no control or inappropriate control treatments such as continuous maize with zero inputs. In assessing whether or not hedgerow intercropping can provide a sustainable and more productive alternative to farmers’ current practices, it is essential that those practices be represented as control treatments within the experimental design. Few farmers grow maize continuously without any inputs at all.
While these acknowledged flaws in design are not peculiar to hedgerow intercropping trials, they certainly add to the complexity of reviewing the literature in order to ascertain under what biophysical conditions hedgerow intercropping will work. This task is already challenging enough given the wide range of tree species, hedgerow management and spacings, crops and local modifications (livestock inclusion, fallow periods) which have been evaluated by scientists throughout sub-Saharan Africa. Added to this complexity are the crucial factors affecting the adoption of this technology by farmers and its long-term impact on their welfare. Several such reviews have been undertaken in recent years and a consensus is emerging concerning the biophysical and socioeconomic conditions under which hedgerow intercropping will benefit farmers (Nair, 1990; Kang, 1993; Rao, 1994; Akyeampong et al., 1995; Sanchez, 1996; Reynolds, 1994; Woomer et al., 1995; Carter, 1995; Whittome, 1994). The biophysical parameters required for success appear to be:
• Soils of moderate pH (greater than 5.5) with a high base status for nutrient recycling in biomass
• Declining soil fertility (recognized as a serious problem by farmers)
• Scarcity of trees in the landscape (thus interest in tree products)
• Rainfall of more than 1000 mm a−1 (adequate for crops and trees)
• Cropping systems dominated by maize (in general, maize responds better to hedgerow intercropping than other crops such as cassava, upland rice, soyabean, and groundnuts)
• Well adapted tree species/provenances for high biomass production
• Timely pruning, efficient utilization of prunings, and sustained hedge growth
• Absence of serious termite attack on the hedge during establishment for certain susceptible species.
Many socioeconomic parameters have also been identified through extensive participatory on-farm research which appear important both in terms of farmer interest and capacity to meet the management requirements (establishment, pruning, and biomass distribution) of the technology. Principal among these are:
• High land pressure associated with declining or absent fallow periods
• Knowledge or tradition of producing seedlings for hedge establishment and filling gaps
• Available labor at pruning time, preferably male as the task is often considered too onerous for women
• Individual secure tenure of discrete parcels of land
• Confinement of livestock to avoid dry season browsing of hedges
• Farm revenue as a major source of household income
• Infrastructural arrangements that promote extension-farmer and farmer-farmer linkages
• A clear perception by farmers that they are substantially benefiting from the technology.
This last point is of overriding importance and is, of course, clearly linked with the biophysical performance of the technology. Two observations have perhaps encapsulated the current status of the acceptance of hedgerow intercropping. First, reviews of long-term and well-designed trials have identified biophysical situations where the technology has failed to work but have been less successful in finding examples where substantial and sustained increases in crop yields have been achieved (Sanchez, 1996). Second, studies of farmers’ reaction to and acceptance of hedgerow intercropping have concluded that, in spite of more than a decade of research, development and promotion, adoption of this technology in sub-Saharan Africa is very low (Fig. 4.6; Carter, 1995; Whittome, 1994; Reynolds, 1994).

Is hedgerow intercropping therefore merely a scientific curiosity, destined to become labeled as one more example of scientists’ “technology push”? Certainly at the moment the evidence is not encouraging, but it still may well be a technology for the future. Why? It is reasonable to imagine that almost all the required socioeconomic criteria identified here will become more conducive to the adoption of hedgerow intercropping as populations continue to increase, agriculture becomes more intensive and farming and its infrastructural support mechanisms become more developed. But what of the essential improvements required in the biophysical performance of the technology? Ong (1994) developed an equation to describe the component effects of hedgerow intercropping, which in its simplest form can be written as:
where I is the overall interaction as a percentage increase in yield achieved over sole crops, free from hedge influence; F the percentage crop yield increase due to fertility improvements (including soil physical improvements and changes in microclimate) from the hedge prunings and roots; and C the percentage decrease in yield due to crop competition with trees for light, water, and nutrients. The challenge is clear: can we maximize F (i.e., increase biomass production and its efficiency as a green manure) and minimize the competition effects (C)?
There are many published examples of contrasting pruning regimes, biomass management strategies and species comparisons, all studied with the objective of optimizing biomass production and its efficiency of use. Very positive results have been reported and yet in many cases the crucial issues of design criteria cast some doubt on the validity of the results. For example, apparently useful modifications to pruning time and biomass management were assessed in a trial at Chitedze Research Station, near Lilongwe in Malawi, which receives an average of 900 mm unimodal rainfall per season (Table 4.1). At the time of the third pruning during the cropping cycle (a time at which the crop is too mature to benefit from additional nutrients), the hedge prunings were sun-dried and stored and then applied as a handful of leaves buried around each plant in the next season 1–2 weeks after maize planting. This practice conserved about two-thirds of the annual biomass produced and utilized it in the most temporally and spatially efficient manner possible. There is plenty of evidence (see Table 4.5) to show that the greatest responses of maize to green manure are achieved when the green manure is incorporated into the soil near the time of maize planting. When this practice was combined with increasingly timely hedge pruning to minimize hedge/tree competition, substantial responses to hedgerow intercropping were achieved in 1991/92, a drought year when competition for moisture was likely to be severe (Table 4.1). It is interesting to note that as the hedge pruning regime improved between 1989 and 1992, the competition effects on the maize adjacent to the hedge declined. It is also impressive that maize yields in the Senna spectabilis hedgerow system were considerably greater than those of maize in the control plot, which received 100 kg N ha−1. However, in this trial the control treatments were separated from the “hedge” treatments by only four guard rows of maize (3.6 m). C. K. Ong (personal communication) has found that in the first year of establishment, senna roots spread 4 m laterally at a semiarid location in Kenya; thus it seems highly probable that they invaded and mined the control plot at Chitedze after 4 years.
Table 4.1
Maize grain yields (t ha−1) in an alley cropping hedge screening trial at Chitedze Research Station, Malawi.
| 1989/90a | 1990/91b | 1991/92c | |
| Sole crop maize | |||
| 0 kg N ha−1 | 1.11 | 0.82 | 0.40 |
| 100 kg N ha−1 | 2.08 | 4.53 | 1.93 |
| Maize + Senna spectabilis hedge | |||
| Rows next to hedge | 0.35 | 0.89 | 2.66 |
| Center rows | 1.11 | 1.02 | 2.48 |
| Mean | 0.73 | 0.96 | 2.57 |
| Maize + Gliricidia sepium hedge | |||
| Rows next to hedge | 1.12 | 1.01 | 1.37 |
| Center rows | 1.48 | 0.96 | 0.98 |
| Mean | 1.30 | 0.99 | 1.18 |

aThe trial was established in the 1987/88 season; in the 1989/90 season there were two primings, the first at 4 weeks after planting (WAP), the second after harvest.
bIn the 1990/91 season, the first pruning was three WAP and the second after harvest.
cIn the 1991/92 season there were three prunings: one WAP, midseason, and after harvest.
Source: From Bundersen (1992)
However, other interesting evidence is also provided by a second trial in Malawi at Makoka Research Station near Zomba (Maghembe, 1994), which confirmed that minimizing tree-crop competition coupled with efficient management of the prunings of spatially mixed trees enhances crop yields. In this trial, Gliricidia sepium was established in a maize crop at a similar density to that commonly used in hedgerows, but at a dispersed spacing of 0.9×1.5 m rather than as a hedge. During the growing season, competition was minimized by cutting back the gliricidia to a height of 10 cm at the time of land preparation in October and twice more during the growing season in December and February. All the biomass was incorporated into the ridges on which the maize was grown (Fig. 4.7). The trial was established in 1992 and the cycle of management was repeated in 1993 and 1994. Root barriers were installed at the start of each season to help eliminate belowground interference between plots.

No major effects were observed in the season of establishment but in the second season, 1993, the maize demonstrated a substantial response to the accumulated additions of green manure, giving 2.5 t grain ha−1 compared with 1.2 t ha−1 from sole maize control plots receiving no N or P and 2.4 t ha−1 from plots receiving 50% of the recommended rates of N and P. In addition, 2.4 t wood ha−1 was produced, currently valued at US$25 t−1. Very similar results were obtained in the third season, 1994. Here is early but solid evidence that minimizing competition and optimizing the efficiency of use of biomass can give positive benefits in spatially mixed systems. This system is now being evaluated with farmers in Zambia.
Other modifications to hedgerow intercropping have also given positive results, in particular the inclusion of short fallow periods during which hedgerows are allowed to grow unchecked and cropping between the hedges is temporarily halted. Reynolds (1994) recently concluded that “alley farming cannot maintain soil fertility without external inputs, or the inclusion of fallow periods.” A good example of this has been observed from a trial, free of known design faults, at Yaoundé in Cameroon which receives a bimodal rainfall of 1600 mm a−1 (Duguma et al., 1994). Mixtures of Leucaena leucocephala and G. sepium were established in 1988 as hedgerows 4 m apart in large plots of 11×16 m. Maize was grown in one season each year only, in both hedgerow plots and control plots of sole unfertilized maize. The hedges were pruned back at the time of maize planting and twice more during the growing season to minimize competition for light. The biomass was incorporated into the soil. In the second season of each year, plots were left either as a tree fallow in hedgerow plots or as a natural fallow in control plots (Table 4.2). The inclusion of a natural fallow period in the control treatment resulted in only a slow decline in maize yield on this moderately fertile Oxisol. However, the inclusion of a tree fallow period in the hedgerow intercropping plot has maintained yields at a high level. Soil analyses (0–15 cm) undertaken in 1994 showed significantly higher organic matter, total N and pH levels in the hedgerow plots. Here again is evidence that a modified hedgerow intercropping system can sustain maize yields at high levels over time. Similar results have been reported by Jabbar et al. (1994) from a bimodal rainfall location (1250 mm a−1) near Ibadan in Nigeria.
Table 4.2
Maize grain yields (t ha−1) in sole maize and hedgerow intercropping plots at Yaoundé, Cameroon.
| 1990 | 1991 | 1992 | 1993 | 1994 | ||||||
| Season | Season | Season | Season | Season | ||||||
| 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | |
| Sole maize | 1.52 | NFa | 2.98 | NF | 3.54 | NF | 2.54 | NF | 2.17 | NF |
| Hedgerow intercropping | 2.13 | TFb | 3.70 | TF | 4.79 | TF | 5.09 | TF | 4.55 | TF |
| SED | 0.38 | – | 0.29 | – | 0.35 | – | 0.44 | – | 0.14 | – |
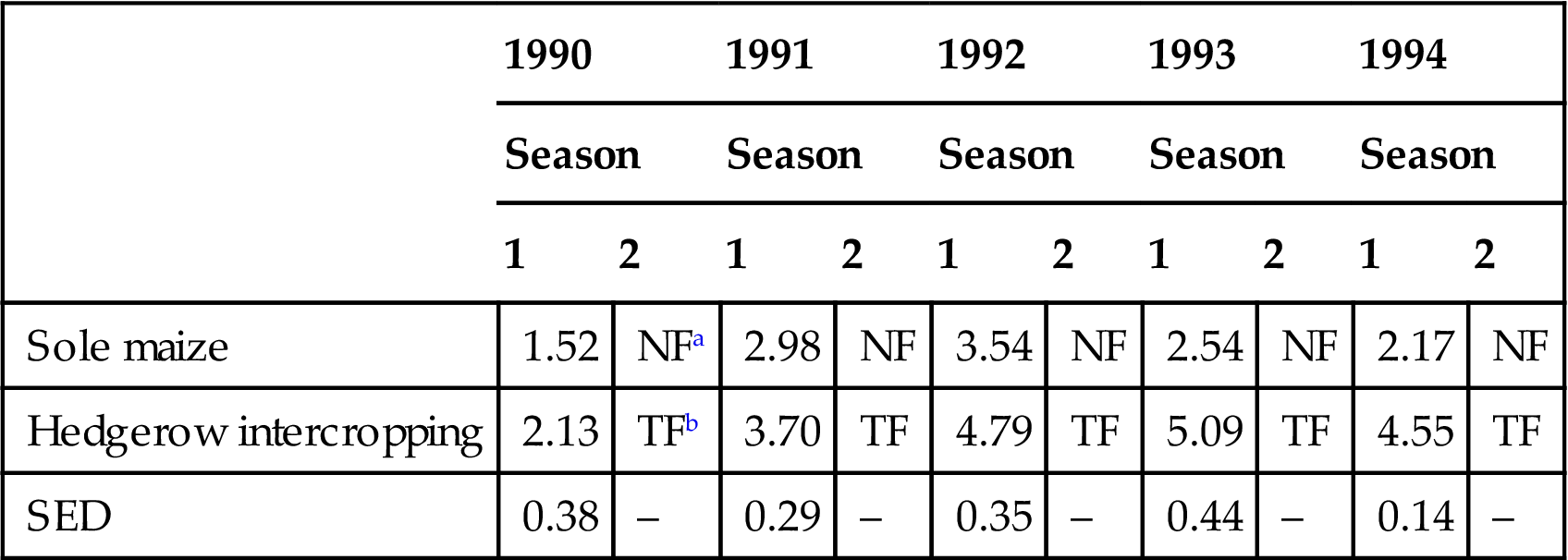
aNF, natural fallow.
bTF, tree fallow.
Source: From Duguma, B., Mollet, M., Tiki Manga, T., 1994. Annual Progress Report. Institute of Agronomic Research (IRA), Cameroon and International Center for Research in Agroforestry (ICRAF) AFRENA Report.
A third success story for hedgerows has been their use in contour plantings on moderately and steeply sloping land for soil erosion control. Here the evidence of benefits is concrete, as discussed further in the section on soil conservation.
In conclusion, as far as hedgerow intercropping is concerned, we suggest that on relatively fertile soils with sufficient rainfall there is emerging evidence that the performance of this technology can be made sufficiently impressive for it to be of interest to farmers. It is important to remember that much of the research conducted on hedgerow intercropping has focused on the nitrogen response of the system. As such, other major plant nutrients were often applied as a blanket dressing. Even under conditions where the system maintains nitrogen levels, it will almost certainly be necessary to add other nutrients, especially phosphorus, for the long-term sustainability of system production.
We also believe that, as agriculture intensifies in such high potential conditions, variants of hedgerow intercropping will become increasingly acceptable through their integration with other systems. In conditions where moisture is limiting, and on inherently infertile and strongly acid soils, the evidence suggests that this is an inappropriate technology.
Improved Fallows
As indicated earlier, trees are a component of naturally regenerating fallows in many traditional shifting cultivation systems throughout Africa and the rest of the tropics. Farmers in many humid tropical regions have recognized certain tree species as being associated with improved soil fertility and therefore deliberately promote such species in fallows either by planting them toward the end of the cropping phase or by selectively controlling those species that compete with the preferred species. Such tree fallows are known as enriched fallows. In southeastern Nigeria farmers encourage species such as Dactyladenia (Acioa) barteri, Alchornea cordifolia, Anthonata macrophylla, Crestis ferrugina, Dialium guineense, and Harungana madagascariensis in the fallow period (Kang et al., 1990). Similarly, farmers in Latin America are known to encourage certain economic and soil improving indigenous trees such as Stryphnodendron excelsum, Dalbergia tucurensis, Dipteryx panamensis, Vochysia ferruginea, and Tabebuia rosea (Montagnini and Sancho, 1990).
With increasing population pressure the inevitable need for shortening the fallow period and increasing cropping intensity has been well recognized. However, 3–6 years of natural tree fallows cannot maintain soil fertility at similar levels to those achieved in traditional shifting cultivation systems (Aweto et al., 1992). Because of this, a concept of improved fallows is now being pursued which involves the planting of fast-growing, nitrogen-fixing, and deep-rooting trees to enhance and maintain soil fertility over a shorter span of time.
Unlike spatial mixtures such as hedgerow intercropping, there is no competition between the tree and the crop component in improved fallow systems (except between adjacent fallow and cropland, which could be important on small farms) as they follow one another in temporal sequence. However, the same principles of experimental design as highlighted in the previous section are still important. Above- and belowground “mining” of resources can still occur in experiments where tree and crop plots (controls) are adjacent. It will also occur when fallows of different lengths are being compared and the fallow treatments are incorrectly phased. For example, crops following a 1-year fallow will suffer interference from adjacent 2- or 3-year fallow plots still in the ground (Rao et al., 1990).
Research on improved fallows in Africa has not been as extensive as that on hedgerow intercropping, but it is gathering momentum and impressive results are being obtained. A good example of such research comes from the Miombo ecozone of southern Africa where shifting cultivation systems such as Chitemene are no longer sustainable. If tree crowns are lopped, 20–30 years’ regeneration is required to sustain the Chitemene type of shifting cultivation in northern Zambia (Mansfield et al., 1975). If trees are coppiced, Stomgaard (1985) estimated that 43 years are required for miombo regeneration. However, recent reports indicate that the commonly practiced fallow period in northern Zambia is now less than 10 years (Kwesiga and Kamau, 1989) and on the plateau in eastern Zambia bush and grass fallows of 1–5 years are common (Kwesiga and Chisumpa, 1990).
Early research with improved fallows at Msekera Research Station at Chipata in Zambia, which receives between 800 and 1000 mm unimodal rainfall, has been encouraging but suffered from design faults (Kwesiga and Coe, 1994). Two-year fallows of Sesbania sesban, planted at a density of 10,000 trees ha−1, resulted in large increases in yields compared with unfertilized maize in the first 2 years after fallow clearance. Yields of 5.0 and 5.6 t ha−1 were obtained in 1990 and 1991 compared with 4.9 and 4.3 and 1.2 and 1.9 t ha−1 from continuously cropped maize with (112 kg N ha−1) and without fertilizer, respectively. In addition, approximately 10 t fuelwood ha−1 were harvested at fallow clearance (Fig. 4.8). However, as already indicated, incorrect phasing in the experimental design and small plot size almost certainly resulted in above- and belowground interference between tree and crop plots in this trial (Kwesiga and Coe, 1994). Research on improved fallows in Zambia has subsequently expanded, with the use of larger plots, root barriers or trenches surrounding plots, and properly phased designs. Furthermore, a wider range of potential fallow species are now being evaluated as 1-, 2- and 3-year fallows.

S. sesban continues to show its superiority as a 1- or 2-year fallow, and Cajanus cajan and Tephrosia vogelii are also showing promise (Table 4.3). These species are currently being jointly evaluated with over 150 farmers in eastern Zambia, and increasingly farmers are beginning to establish spontaneously their own fallow experiments with sesbania. Farmers who have already harvested their first crop of maize after fallow clearance are observing similar yield increases to those reported in Table 4.3 (Fig. 4.9). Similar maize responses have been observed on acid Ultisols in Onne, Nigeria (Gichuru, 1991) where a 2-year fallow of Tephrosia candida gave maize yields of 2.46 t ha−1 compared with 0.95 t ha−1 following a natural bush fallow. On the basis of a survey of farms under G. sepium fallow in southern Nigeria, Adejuwon and Adesina (1990) concluded that soil fertility improved progressively with the length of fallow. The most significant improvements were in terms of organic matter, nitrate-nitrogen, and potassium. Unfortunately, no test crop data from this study are available.
Table 4.3
The effect of 1- and 2-year fallows of different species on maize yield in the first year after clearance at Chipata, Zambia, 1994.
| Maize yield (t ha−1) | ||
| Grain | Stover | |
| Sesbania sesban | ||
| 1-year fallow | 3.43 | 4.60 |
| 2-year fallow | 5.47 | 7.01 |
| Tephrosia vogelii | ||
| 1-year fallow | 2.80 | 3.49 |
| 2-year fallow | 3.65 | 5.34 |
| Sesbania macrantha | ||
| 1-year fallow | 2.08 | 4.56 |
| 2-year fallow | 3.00 | 4.22 |
| Cajanus cajan | ||
| 1-year fallow | 2.40 | 4.05 |
| 2-year fallow | 2.78 | 4.39 |
| Grass fallow | ||
| 2-year fallow | 2.12 | 3.69 |
| Continuous maize | ||
| With fertilizer | 3.40 | 5.09 |
| Without fertilizer | 1.09 | 2.33 |
| Groundnut/maize rotation | 2.21 | 3.52 |
| SED | 0.44 | 0.63 |


Such improved fallows clearly work and, as is the case in Zambia, farmers are interested and wish to try them. Initially it was thought that this practice would be most relevant to farming systems where land is not a constraint and farmers are setting, or could afford to set, land aside for fallowing. However, recent surveys in western Kenya, in a high population density zone (300–1000 people km−1 with a bimodal rainfall of 1800 mm a−1, have shown that nearly 50% of farmers with farms of between 0.5 and 5.0 ha leave their land under fallow for one or two seasons). Severe land depletion, with maize yields falling below 0.5 t ha−1, and an inability to purchase inputs, are the principal causes, but insufficient labor to cultivate crops was also cited as a reason (ICRAF, 1994, p. 27). These farmers have expressed interest in short-duration improved fallows for both soil fertility improvement and wood production.
In the Shire highlands of Malawi, a unimodal rainfall zone, farm sizes are even smaller, ranging from 0.2–0.5 ha, and preclude the setting aside of land for fallow. As expected, soil fertility is declining, with many farmers unable to afford fertilizer. Many farmers currently intercrop (or relay crop) their maize with C. cajan, both as a source of income from the local sale of seed and also as a means of adding organic nitrogen through above- and belowground litter. Cajanus is known to fix about 90% of its nitrogen requirements from the atmosphere (Kumar Rao et al., 1987) and other studies (Poth et al., 1986) have shown residual benefits to subsequent maize crops equivalent to 40 kg N ha−1. An innovative form of dry season fallow (relay cropping) is being investigated in which no land is taken out of maize production. S. sesban is established as seedlings at the same time as maize, which is planted and managed according to the recommended practice. Sesbania is planted at a spacing of 90×150 cm and maize at 30×75 cm. Superimposed are three levels of fertilizer application to the maize, 0%, 50% and 100% of the recommended amount for Malawi (14 kg N ha−1 and 10 kg P ha−1 at planting, plus 42 kg N ha−1 as a top dressing) (ICRAF, 1995).
After the harvest of maize, the sesbania is allowed to continue growing throughout the dry season on residual moisture (Fig. 4.10). Prior to the sowing of maize in the rainy season of the following year, the sesbania is harvested and the leaves, pods, and small branches incorporated into the soil. Material suitable for fuelwood is separated and removed. This cycle has been repeated on the same plots for five consecutive seasons (Table 4.4).

Table 4.4
Maize yields (t ha−1) obtained in a relay cropping system with Sesbania sesban at Makoka, Malawi.
| Fertilizer (% of recommended amounts for Malawi) | |||||||
| 0 | 50 | 100 | |||||
| Sole maize | Relay cropping | Sole maize | Relay cropping | Sole maize | Relay cropping | SEDa | |
| 1989/90 | 1.00 | 1.02 | 3.75 | 3.27 | 4.48 | 3.77 | 0.40 |
| 1990/91 | 1.33 | 2.41* | 6.21 | 6.13 | 7.12 | 7.84 | 0.60 |
| 1991/92 | 0.67 | 2.57** | 2.99 | 3.75* | 3.47 | 4.32 | 0.38 |
| 1992/93 | 1.02 | 2.33** | 4.79 | 5.99* | 6.06 | 7.40** | 0.51 |
| 1993/94 | 0.51 | 1.16* | 2.41 | 3.29** | 3.23 | 3.49 | 0.36 |
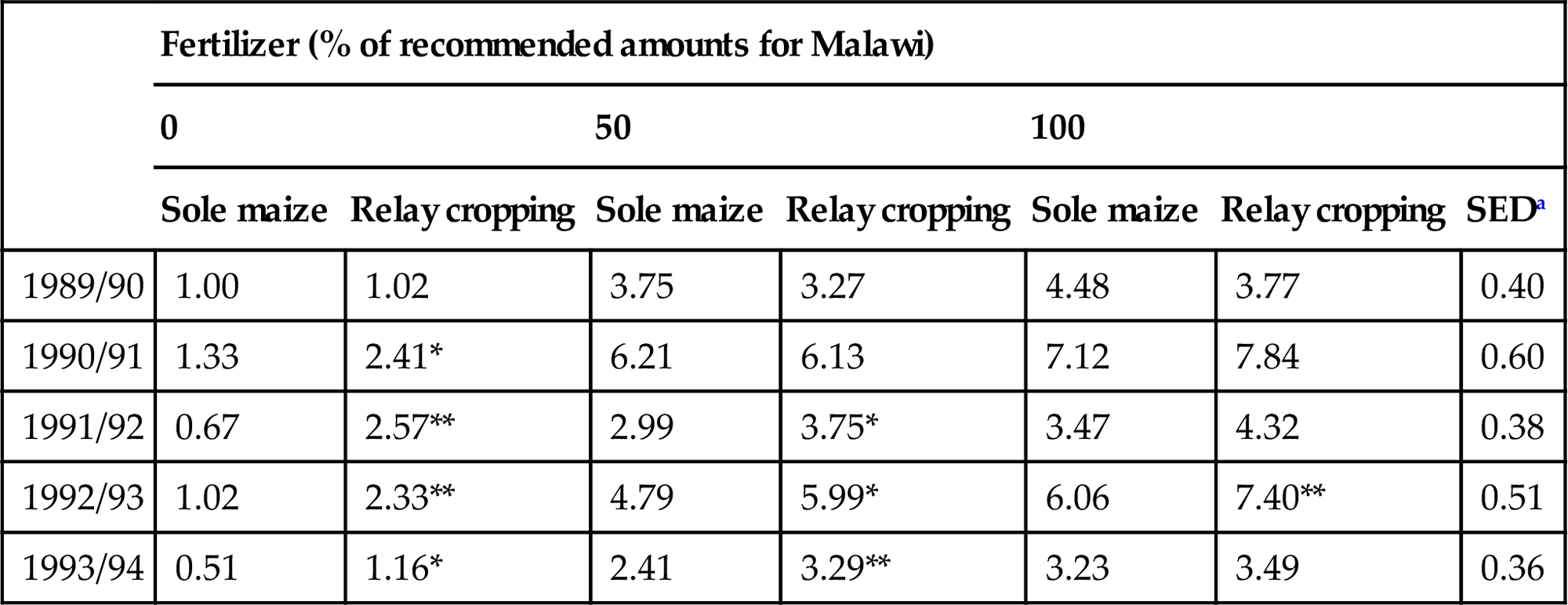
*Significant difference at 10%. **Significant difference at 5%.
aSED provided is for comparison of yield differences between sole and relay cropped maize within fertilizer levels.
Source: From Maghembe, J.A., 1994. Out of the forest: indigenous fruit trees in southern Africa. Agrofor. Today 6 (2), 4–6.
In the season of establishment (1989/90), as expected, the relay cropped plots within any given fertilizer subtreatment showed no benefit of the technology, although significant responses to fertilizer were found and continued to be observed throughout the life of the trial. In the second season, the effect of incorporating sesbania residue into the soil prior to maize sowing gave significant increases in the unfertilized maize subtreatment, and this effect continued to be observed in subsequent seasons, both in the unfertilized maize crop and in maize receiving 50% of the recommended rate of fertilizer. A similar trend was also observed in the maize receiving 100% of the recommended rate of fertilizer but the effect was only significant in the climatically favorable season of 1992/93, when rainfall was high and well distributed.
Fuelwood is in very short supply in the Shire Highlands and is currently valued at US$25 t−1. The added benefit of the fuelwood produced from the dry season sesbania fallow (mean of 2.08 t ha−1 across treatments and years) has aroused considerable interest among farmers who have seen the performance of this technology. Again, however, there is some concern that the small plot size used in this experiment may have resulted in unrepresentative exploitation of the belowground soil resources during the dry season. Thus in the 1994/95 season, relay cropping with S. sesban is being evaluated in large plots directly with farmers in the Zomba district.
In conclusion, with respect to improved fallows, various forms appear to have considerable potential as a strategy to restore soil fertility and improve crop yields, and, as far as farmers are concerned, do not suffer the complexity of managing tree-crop competition. In addition, short-term fallows have the potential to produce useful quantities of firewood which appears to be attractive for farmers in many areas (see Fig. 4.1). Although yet to be proven, the technology may be widely acceptable since it builds on a widespread tradition and knowledge of fallowing as currently practiced by farmers when crop yields fall below a certain economic threshold.
Appropriate fallow tree species have been identified for a wide range of soil and climatic conditions (Rao, 1994). This is important because care must be taken not to overpromote a single species, such as S. sesban, which is known to be susceptible to attack by the Mesoplatys beetle. In addition, sesbania can lead to a build-up of soil nematode populations, so is unsuitable for use prior to susceptible cash crops such as tobacco.
Research on improved fallows is likely to expand in coming years and it is essential that new experiments pay careful attention to experimental design issues (Rao et al., 1990; Coe, 1994) in order to avoid the pitfalls which may have befallen much of the previous research on hedgerow intercropping. It is also important that long-term trials be established to assess how sustainable such improved fallows are over a longer period. Nitrogen-fixing trees will certainly maintain soil nitrogen at high levels, but other major plant nutrients such as phosphorus may well become limiting after several cycles of fallowing.
As with any intervention, the adoption of improved fallows will depend upon farmers’ perception of the benefits, particularly when additional labor is required in fallow establishment and management. Clearly, farmers recognize an economic threshold below which they do not consider cropping to be worthwhile and this is the underlying cause for land abandonment and natural fallows. Research must demonstrate that the returns to labor for improved fallows are superior to those achieved through natural fallows. Such analysis (ICRAF, 1994, p. 111–114) demonstrated substantially greater returns to labor when a 2-year S. sesban fallow was compared with continuous maize cultivation over a 6-year period. Data for a more appropriate comparison with a natural fallow were not available in the data set studied. Long-term trials with appropriate controls are required to address this issue but ultimately it will be the farmers who decide. For this reason, participatory research with farmers, as is currently being undertaken in Zambia, will be the most relevant approach.
Biomass Transfer
The transfer of tree or shrub biomass from distant areas to food crop production fields to maintain soil fertility and crop yields is a well-known practice in certain countries. For instance, such a practice has been used for decades in Asia for irrigated rice production. Gliricidia maculata is grown on the paddy field bunds and incorporated into the soil before rice transplanting (Singh et al., 1991). In southern Africa, collection and use of miombo woodland litter as a source of plant nutrients is a common practice. Farmers use litter directly as green manure or they dry and store it for later use, especially if the material is collected during the noncropping period when the demand for labor is low. In addition, it may be cured in cattle pens and mixed with manure (Nyathi and Campbell, 1993). The traditional Chitemene and fundikila shifting cultivation systems in northern Zambia and its surroundings also heavily depend on exploiting miombo litter (Matthews et al., 1992). Recent research at Domboshawa in Zimbabwe has demonstrated that high-quality litter gives substantially greater maize yields than miombo leaf litter (Dzowela et al., 1994). Yields of 5.7 and 5.6 t ha−1 were obtained following the incorporation of 5 t litter ha−1 from L. leucocephala and C. cajan, respectively, at planting compared with only 2.5 t ha−1 achieved through the incorporation of miombo litter at the same rate.
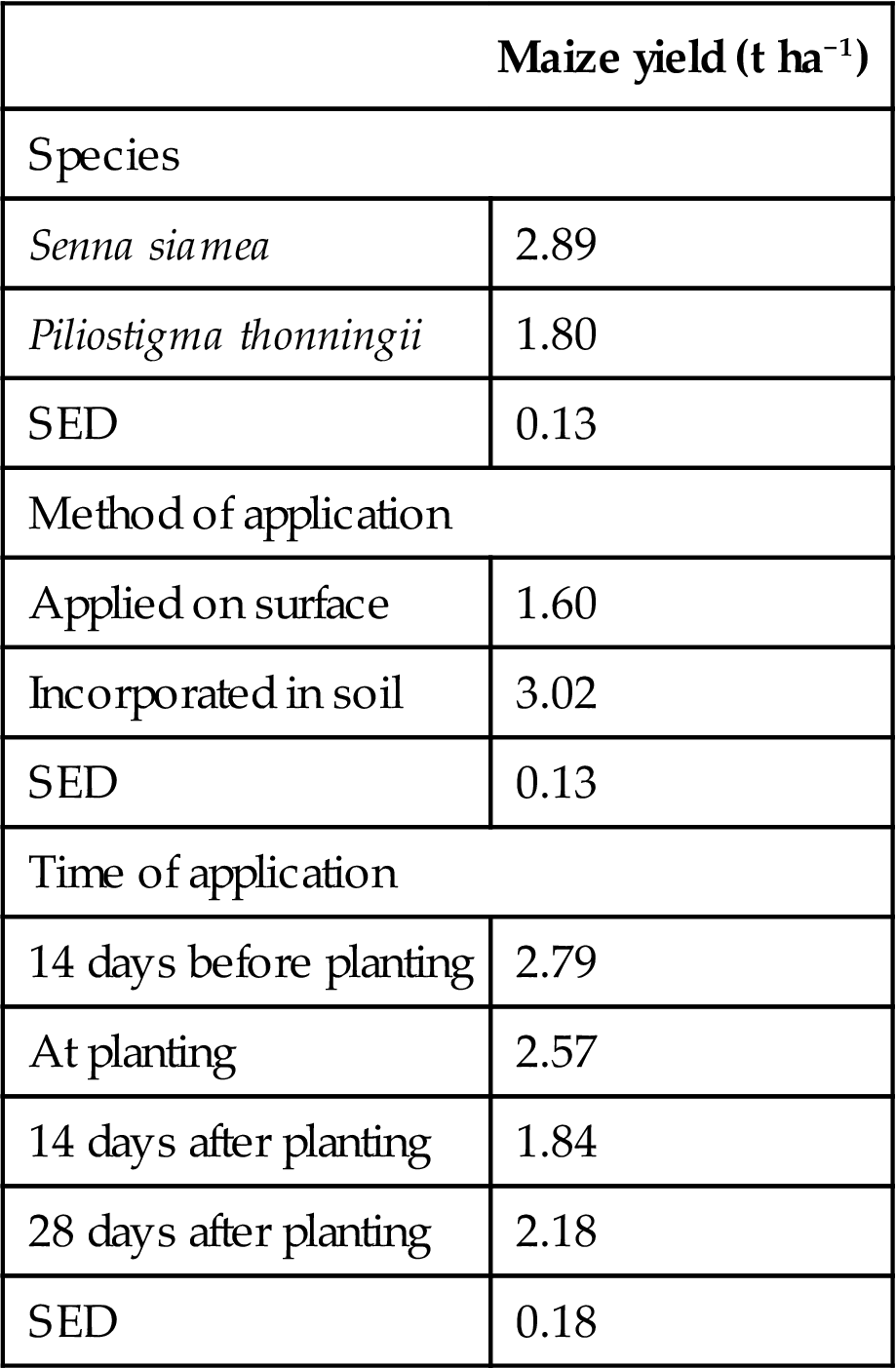
Several factors affect the response of crops to the application of transferred biomass and have been reviewed by Rao (1994). Major factors are the chemical composition of the litter of different species and the method and timing of application. The impact of these factors is illustrated by results obtained in Zambia (Table 4.5). These results confirm that incorporation of the litter into the soil close to the time of maize planting produces the greatest maize yield response. However, with species that decompose rapidly (such as gliricidia and sesbania) the timing is less critical and incorporation may be delayed for 3 or 4 weeks after planting (Read et al., 1985). Palm (1996), from an extensive review of the literature, concluded that, as a general guideline, if the concentration of nitrogen in the green manure is less than 1.74%, then net nitrogen immobilization will occur and will possibly continue for the bulk of the cropping season. If the concentration of nitrogen is greater than 1.74%, then net mineralization will occur, but will decrease as a function of the ratio of lignins+polyphenols to nitrogen.
Table 4.5
The effect of species, method of application and time of application of mulch on maize yields at Chipata, Zambia, 1993 (main effects from a factorial trial).
| Maize yield (t ha−1) | |
| Species | |
| Senna siamea | 2.89 |
| Piliostigma thonningii | 1.80 |
| SED | 0.13 |
| Method of application | |
| Applied on surface | 1.60 |
| Incorporated in soil | 3.02 |
| SED | 0.13 |
| Time of application | |
| 14 days before planting | 2.79 |
| At planting | 2.57 |
| 14 days after planting | 1.84 |
| 28 days after planting | 2.18 |
| SED | 0.18 |
Source: From ICRAF, 1994. Annual Report 1993. International Center for Research in Agroforestry, Nairobi, Kenya, p. 118.
Many studies have, however, confirmed that the efficiency of organic nitrogen is lower than that of inorganic nitrogen (Gutteridge, 1992; Read et al., 1985). Several reasons seem to be evident, such as slow decomposition of residues, lack of synchrony in the mineralization of nutrients and crop uptake, unfavorable soil conditions (moisture, temperature, biological activity), and loss of nutrients through leaching, volatilization, and immobilization (Palm, 1996). The need for synchrony between nutrient mineralization and crop demand increases with rainfall and leaching intensity and shallow-rooted crops, especially on acid soils in the humid tropics (van Noordwijk et al., 1991). The less efficient use of organic nitrogen is typified by results obtained from a trial conducted over a 4-year period at Chipata in Zambia. Five rates of inorganic nitrogen (0, 30, 60, 90, and 120 kg ha−1) and organic nitrogen from G. sepium or L. leucocephala prunings (0, 5, 10, 15, and 20 t fresh weight ha−1) were applied in factorial combination on the same plots for four seasons. Basal dressings of phosphorus and potassium were supplied to maize each year to focus on the nitrogen response. Consistent and linear responses of the maize to the different rates of both sources of nitrogen were observed in all seasons, with the response in any given season always being greater to inorganic nitrogen. The results are typified by the regression equations obtained in the 1989/90 season (ICRAF, 1994, pp. 114–117). For gliricidia:
and for leucaena:
where Y is maize yield (kg ha−1) and Ni and No are kg added nitrogen ha−1 in inorganic and organic form, respectively.
These results confirm that nitrogen from gliricidia litter is used more efficiently than that from leucaena. Several researchers have shown that nitrogen release from gliricidia is rapid compared with leucaena (e.g., Constantinides and Fownes, 1994).
However, we must emphasize that in this trial, as in many others, the response to the different sources of nitrogen application was measured on an annual basis. There is a need to examine residual effects in more detail to assess the longer-term relative efficiencies of organic and inorganic nitrogen. It has also been observed that the combination of organic and inorganic nitrogen fertilizers improves the efficiency of nitrogen uptake. Soil microbial activity may be increased and nitrogen capture by the microbial pool from inorganic sources may be improved when fresh organic matter, which can supply soluble carbon as a source of energy, is used. This would reduce leaching and volatization losses, and enhance nitrogen cycling rates and thus nitrogen availability (Buchanan and King, 1992; Snapp, 1995).
There are several other key research issues that remain to be addressed. First, how feasible will it be to find suitable niches on-farm to establish high-quality litter banks? A rough generalization is that approximately 0.5 ha litter bank of an adapted tree species would be required to produce useful amounts of biomass (around 5 t dry matter ha−1) for 1.0 ha of cropped land. Second, how sustainable would a litter bank be before nutrient mining of the soil reduced its productivity? Last, the labor requirements and the opportunity cost of the land set aside for litter banks need to be carefully assessed. Research in Zimbabwe and Kenya (Dzowela et al., 1994; A. Niang, ICRAF, personal communication) is currently addressing these issues.
Soil Conservation
As populations expand, both in numbers and in terms of area settled, more marginal and sloping land inevitably comes under crop production. The consequence of farming fragile soils on sloping land in areas receiving intense rainfall is as inevitable as it has been damaging in the humid and subhumid tropics of Africa. Even in 1938 Hailey concluded that water erosion of soils “is now one of the most serious problems of Africa.” Nearly 60 years later examples of the impact of Hailey’s timely warning can still be seen in the widespread existence of contour bunds in Zimbabwe, the commonly practiced contour ridging in Malawi and the bench terraces of southwestern Uganda (Fig. 4.11). Since that time, researchers and farmers have been acutely aware of the continuing gravity of the problem (Young, 1989; Kiepe and Rao, 1994) and yet mechanical erosion control measures, involving some form of earth movement, are seldom adopted by small scale farmers. Without incentives, as often provided in the developed world, farmers do not perceive the short-term benefits achieved as being worth the cost of installation and maintenance (Mwakalogho, 1986). Young (1989), after an extensive review, concluded that “conservation is likely to be most effective where it is conducted with the active cooperation of farmers, in their perceived interests, and integrated with other measures for agricultural improvement.” Agroforestry can achieve this through its capacity to combine short-term production with longer-term soil conservation functions.

Three broadly defined agroforestry systems have potential in the humid and subhumid tropics of Africa: barrier hedges planted on the contour, tree/shrub combinations to stabilize existing conservation structures and multistrata systems that simulate natural forests and woodlands. In the semiarid tropics of Africa, linear plantings of trees as windbreaks have shown considerable potential to reduce wind erosion but fall outside the scope of this review. Their potential is reviewed elsewhere (FAO, 1986; Kiepe and Rao, 1994).
Barrier Hedges
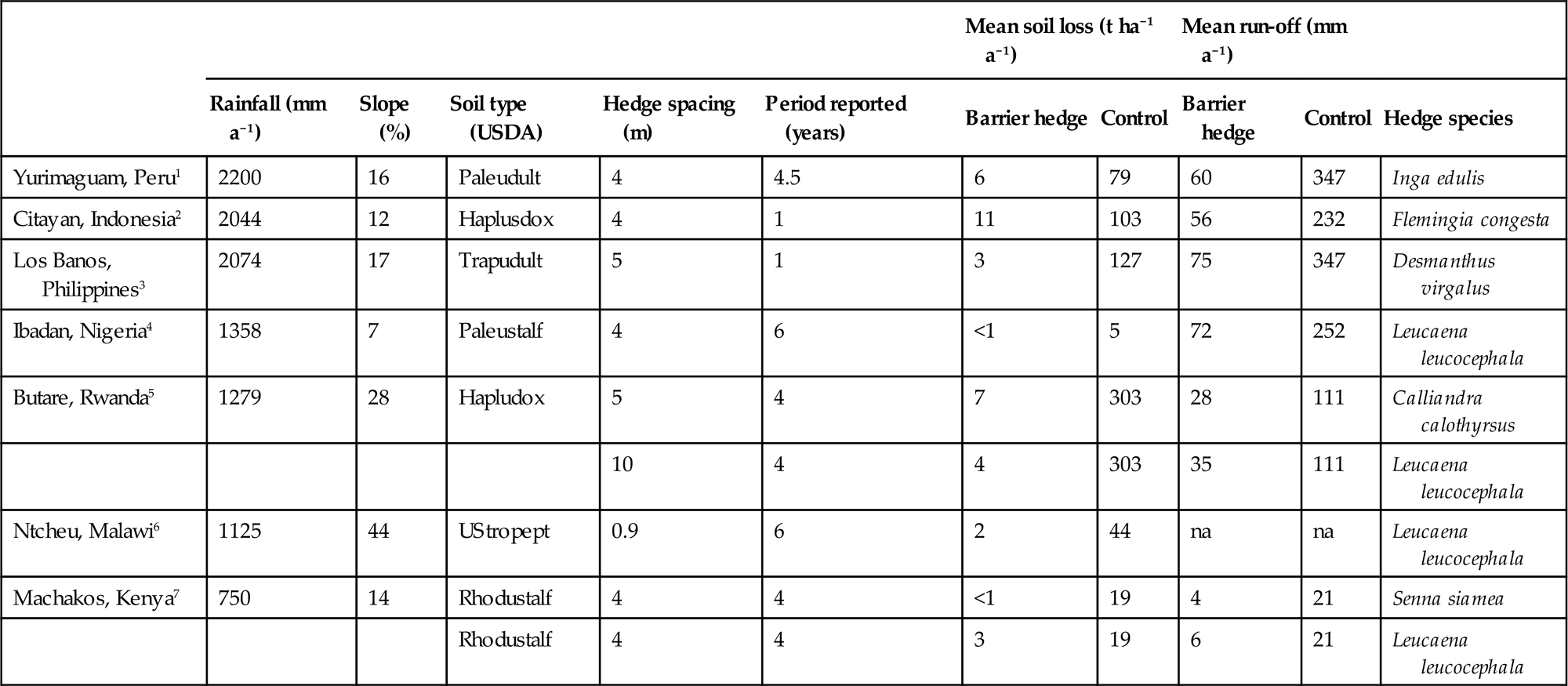
Barrier hedges, planted on the contour line, have been widely evaluated throughout the humid and subhumid tropics. Optimum spacing between hedges is dependent on the degree of slope but within-hedge spacing is commonly between 10 and 25 cm. Hedges are pruned frequently to minimize competition effects with adjacent crops in a similar manner to hedgerow intercropping. Since erosion control is the principal objective, hedgerow prunings are usually applied as a surface mulch to provide added soil protection, a factor which can be critical during the early growth stages of crops. Long-term and properly designed trials, in which surface water run-off and soil erosion losses from land under barrier hedges are actually recorded, are not frequently reported in the literature, but the results that are available are encouraging and often dramatic (Table 4.6). Hedges spaced 4–5 m apart appear effective at controlling erosion on moderately sloping land (10–25%) regardless of the annual rainfall or soil type. Other results, however (Anecksamphant et al., 1990; Nyamulinda, 1991), have shown that on severely sloping land (35–60%) in Thailand and Rwanda such a spacing is too great to control soil loss effectively. In contrast, on a slope of 44% at Ntcheu in Malawi Banda et al. (1994) demonstrated that a spacing of 0.9 m almost completely eliminated soil loss (Table 4.6) and also maintained maize yields at about 2 t ha−1 compared with the control where yields declined to 0.2 t ha−1 over 6 years.
Table 4.6
Soil loss and surface water run-off from land under barrier.
| Mean soil loss (t ha−1 a−1) | Mean run-off (mm a−1) | |||||||||
| Rainfall (mm a−1) | Slope (%) | Soil type (USDA) | Hedge spacing (m) | Period reported (years) | Barrier hedge | Control | Barrier hedge | Control | Hedge species | |
| Yurimaguam, Peru1 | 2200 | 16 | Paleudult | 4 | 4.5 | 6 | 79 | 60 | 347 | Inga edulis |
| Citayan, Indonesia2 | 2044 | 12 | Haplusdox | 4 | 1 | 11 | 103 | 56 | 232 | Flemingia congesta |
| Los Banos, Philippines3 | 2074 | 17 | Trapudult | 5 | 1 | 3 | 127 | 75 | 347 | Desmanthus virgalus |
| Ibadan, Nigeria4 | 1358 | 7 | Paleustalf | 4 | 6 | <1 | 5 | 72 | 252 | Leucaena leucocephala |
| Butare, Rwanda5 | 1279 | 28 | Hapludox | 5 | 4 | 7 | 303 | 28 | 111 | Calliandra calothyrsus |
| 10 | 4 | 4 | 303 | 35 | 111 | Leucaena leucocephala | ||||
| Ntcheu, Malawi6 | 1125 | 44 | UStropept | 0.9 | 6 | 2 | 44 | na | na | Leucaena leucocephala |
| Machakos, Kenya7 | 750 | 14 | Rhodustalf | 4 | 4 | <1 | 19 | 4 | 21 | Senna siamea |
| Rhodustalf | 4 | 4 | 3 | 19 | 6 | 21 | Leucaena leucocephala | |||
Source: Adapted from Njoroge, M., Rao, M.R., 1994. Barrier hedgerow intercropping for soil and water conservation on sloping lands. Paper Presented at the 8th International Soil Conservation Organization Conference. Soil and Water Conservation: Challenges and Opportunities. 4–8 December, 1994, New Delhi, India. From 1ICRAF, 1994. Annual Report 1993. Nairobi, Kenya: International Center for Research in Agroforestry, 2Hawkins et al. (1990), 3Paningbatan (1990), 4Lal (1989), 5Konig (1992), 6Banda et al. (1994), 7ICRAF, 1993. ICRAF: The Way Ahead. Strategic Plan. ICRAF, Nairobi, Kenya.
However, this dramatic impact of erosion control on crop yields is not noted in the other trials reported in Table 4.6. Njoroge and Rao (1994) concluded that short-term yield increases cannot be expected where the soils are deep and fertile and soil erosion losses are only moderately high. Positive crop responses are more common in the short term on steep slopes with shallow soils which experience high-rainfall and where soil loss and hence nutrient depletion rates are rapid.
Experiments conducted in the Philippines and Indonesia through the Sloping Lands Network of the International Board for Soil Research and Management (IBSRAM) demonstrated substantial declines in crop yields under farmers’ management where no soil conservation was practiced. In contrast, barrier hedgerows maintained yields at about twice those achieved by farmers’ conventional practice over a 3-year period (IBSRAM, 1994).
In many instances soil loss measurements, such as those reported in Table 4.6, have not been recorded but evidence for soil conservation is inferred through the rapid accumulation of soil on the upslope side of barrier hedges, resulting in the natural formation of “biological terraces” (Fig. 4.12). For example, a drop of between 0.5 and 0.6 m in soil levels between the upslope and downslope side of barrier hedges was reported in the Philippines on slopes of 20–30% after only 3 years (Maclean et al., 1992; Garrity, 1994). Such trapping of soil and the progressive formation of terraces result in the reduction of slope length and angle. Although the natural formation of “biological terraces” is a positive attribute of barrier hedges, it occurs because soil movement is still taking place between hedgerows. The displacement of soil and nutrients from the upper parts of the alley, and their deposition on the lower portion results in rapid formation of a soil fertility gradient across the terrace. Many researchers (Bannister and Nair, 1990; Maclean et al., 1992, Solera, 1993; Garrity, 1994) have observed the rapid development of this phenomenon, and an associated gradient of crop yields. Typically, crop yields are 50–75% less on the scoured upper parts of the terrace than on the lower parts. Such “terrace scouring” is not restricted to barrier hedges. In southwest Uganda, near Kabale, a farm survey was undertaken of sorghum yields across bench terraces constructed 30 years ago on slopes of around 60%. It was observed that terrace scouring was widespread and, averaged across all farms surveyed, sorghum yields were 0.75, 1.10, and 2.85 t ha−1 for the upper, middle, and lower parts of the terrace, respectively (ICRAF, 1994, p. 101) (Fig. 4.11). Little research has been reported that examines ways to prevent or cure the impact of such terrace scouring. Some work is underway in the Philippines which suggests that a combination of contour ridge land preparation and barrier hedges may be effective as a preventative measure (ICRAF, 1995) and early results from southwest Uganda indicate that leguminous trees, planted as fodder banks or woodlots, could play an important role in reclaiming the upper portions of scoured terraces (ICRAF, 1994, pp. 104–106). However, the issue is largely underresearched and remains a priority for future research (Sanchez, 1996).
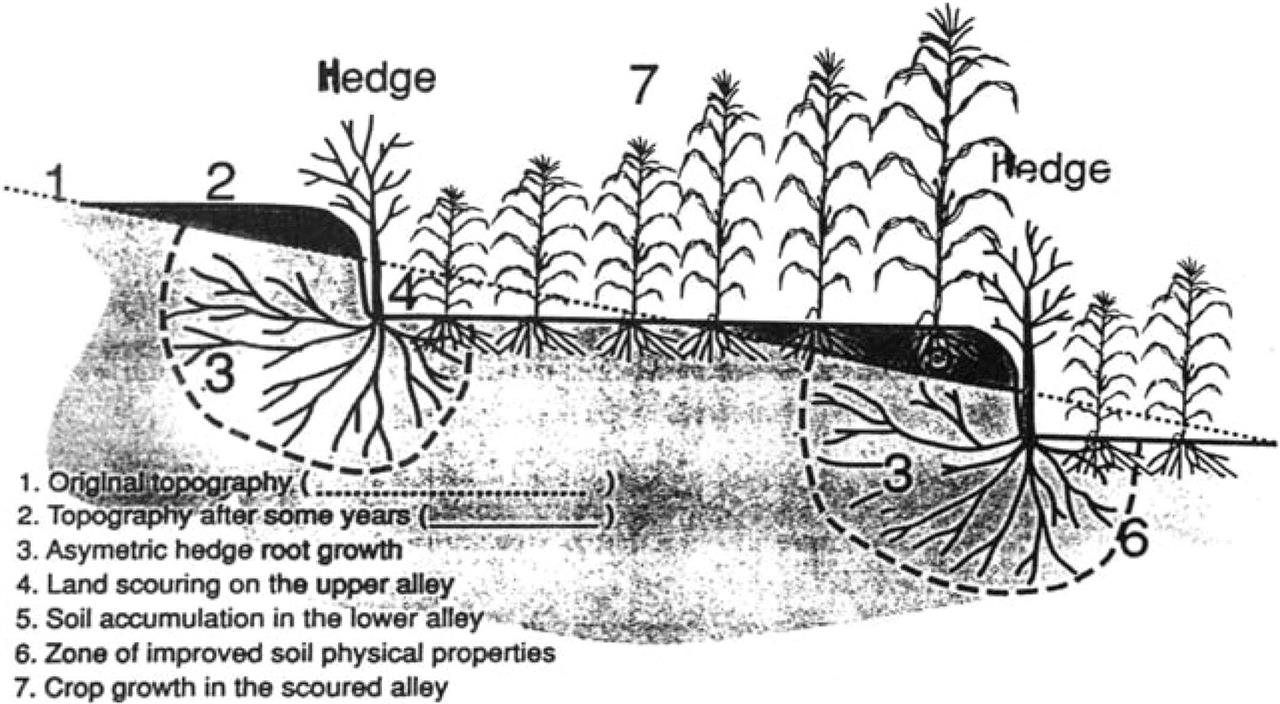
Little detailed research has been undertaken of the processes leading to the effectiveness of barrier hedges but from what evidence is available. Njoroge and Rao (1994) concluded that several probable mechanisms are involved: hedgerows are semipermeable and allow the disposal of excess water without ponding and stagnation; hedge roots stabilize terraces as they form, and improve infiltration rates immediately adjacent to the hedge; surface mulch, when applied, reduces rainfall impact and soil erodability, improves soil fertility and soil physical conditions, and also conserves moisture by reducing soil evaporative loss during early crop growth (Fig. 4.12).
Barrier hedges are not, however, a new concept and have been adopted by farmers in several parts of the world. Kerkhoven first advocated their use in 1913 and widespread planting occurred in the 1930s and 1940s in Indonesia (Metzner, 1976), and more recently in the Philippines and Haiti (Tacio, 1991; Pelleck, 1992) (Fig. 4.13). Currently, adoption in sub-Saharan Africa is low, although the practice is widely recognized as being effective, and is being actively promoted by several development schemes, e.g., in Malawi (Franks, 1992).

In recent adoption studies in the Philippines it was found that farmers had modified the technology by replacing shrubs for mulch production with trees and shrubs with a cash value (Fujisaka, 1993). Analogous to this is the widespread practice of contour planting fodder grasses in Rwanda (Niang et al., 1996), Kenya (O’Neill and Muriithi, 1994), and Burundi (Akyeampong, 1996). The message from farmers seems clear: barrier hedges are an acceptable innovation but in the absence of incentives or short-term crop yield advantages they must produce a valued product rather than green manure. This is corroborated by an economic analysis of alternative uses of leucaena hedge prunings (ICRAF, 1993, pp. 27–29). Prunings used as a fodder supplement for milk production gave economic returns between three and seven times those achieved when they were used as green manure on maize.
However, the use of hedge prunings as fodder, or the use of alternative high-value trees or shrubs, begs the question as to what extent erosion control will be reduced if the surface mulch is not applied to the soil between the hedges. Under mildly erosive conditions at Machakos, Kenya, it was found that, when the mulch from Senna siamea barrier hedges was not applied, the hedges alone reduced annual soil erosion losses from 36 t ha−1 in the control plots to 2 t ha−1 in 1990, and from 12 t ha−1 to 1.5 t ha−1 in 1992 (ICRAF, 1993, p. 63). However, in the same trial in 1994, a highly erosive rainfall of 115 mm occurred in a 24-h period and in this case erosion was only reduced from 46 t ha−1 in the control to 17 t ha−1 in the “hedge alone” treatment. In the “hedge plus mulch” treatment, soil losses were only 1 t ha−1 (C. K. Ong, ICRAF, personal communication). As far as we are aware, studies such as this one are rare. The results indicate that under mildly erosive conditions barrier hedges alone are effective but that, as rainfall intensity (or slope) increases, the added benefit of surface mulching becomes important. However, as noted earlier, it is under such conditions that farmers are likely to observe short-term crop yield benefits (IBSRAM, 1994).
An innovative approach comprising a modification of barrier hedges is being assessed in Mindanao, Philippines, which may well have potential in sub-Saharan Africa (ICRAF, 1994, pp. 140–141). Narrow contour strips of the field area are left uncultivated and allowed to vegetate naturally. The natural vegetative strips capture run-off sediment and form biological terraces in a similar manner to barrier hedges. Farmers in the Philippines have indicated that this low investment form of erosion control is attractive, and that the biological terraces can provide the foundation for agroforestry. They are planting income-generating perennials and fodder species along the terrace risers.
In conclusion, barrier hedges have proved to be effective in controlling soil erosion on gentle to moderate slopes. Under such conditions, hedges alone are sufficient. The adoption of this technology in sub-Saharan Africa could well be enhanced if the hedgerow prunings of leguminous shrubs were used by farmers as a high-quality fodder supplement (see subsequent section) rather than as a surface mulch. Under conditions of more severe soil erosion, the added soil protection of the surface mulch appears important but short-term crop yield benefits which are likely to accrue under such conditions may be a sufficient incentive to farmers. Alternatively, even on steep slopes the application of surface mulch may be avoided if other proven soil conservation practices such as contour ridging and the surface application of crop residues are utilized (Young, 1989, pp. 35–39). Even though such practices will control soil loss from the slope as a whole, it appears that soil movement between barrier hedges is still likely to occur, leading to the terrace scouring effect. More research is required to find ways to prevent this happening and to reclaim degraded soils where scouring has already occurred.
Trees and Shrubs on Conservation Structures
Where conservation structures such as earth bunds, terraces, or conservation grass strips are already in place, trees and shrubs can play an important role in both stabilizing such structures and increasing the diversity of this potential farm niche through the production of fodder, wood, or fruits. Although such niches usually occupy only a small proportion of the land area, they are often fertile due to the accumulation of nutrient-rich soil from terrace scouring.
The inclusion of trees and shrubs on conservation structures is not an erosion control research issue as such since it is the conservation structure itself that is the main focus of research interest. There are nevertheless important research issues which need to be considered. To exploit fully the productivity of conservation structures, the priority needs of farmers must be assessed and potential markets for high-value products ascertained before choices are made from a wide range of possible trees. In identifying appropriate species, spacing and establishment techniques must be known. For example, timber-producing trees with a tall and linear growth habit (i.e., Grevillea robusta, Markhamia lutea, and Alnus acuminata) may be spaced 3–5 m apart but fruit trees such as citrus, mango, avocado, and jackfruit require a spacing of not less than 10 m (Kiepe and Rao, 1994). Where fodder production is a high priority, species such as Calliandra calothyrsus and L. leucocephala (Akyeampong, 1996; Niang et al., 1996a) can be established as closely spaced pure hedges or in combination with existing grass strips. In many instances, farmers may wish to plant a mixture of upper-story and understory trees to provide a range of products. For example, mixtures of upper-story trees for poles (Casuarina equisetifolia and G. robusta) and understory fodder trees (C. calothyrsus) are being evaluated on terrace risers at Kachwakano in southwest Uganda by ICRAF.
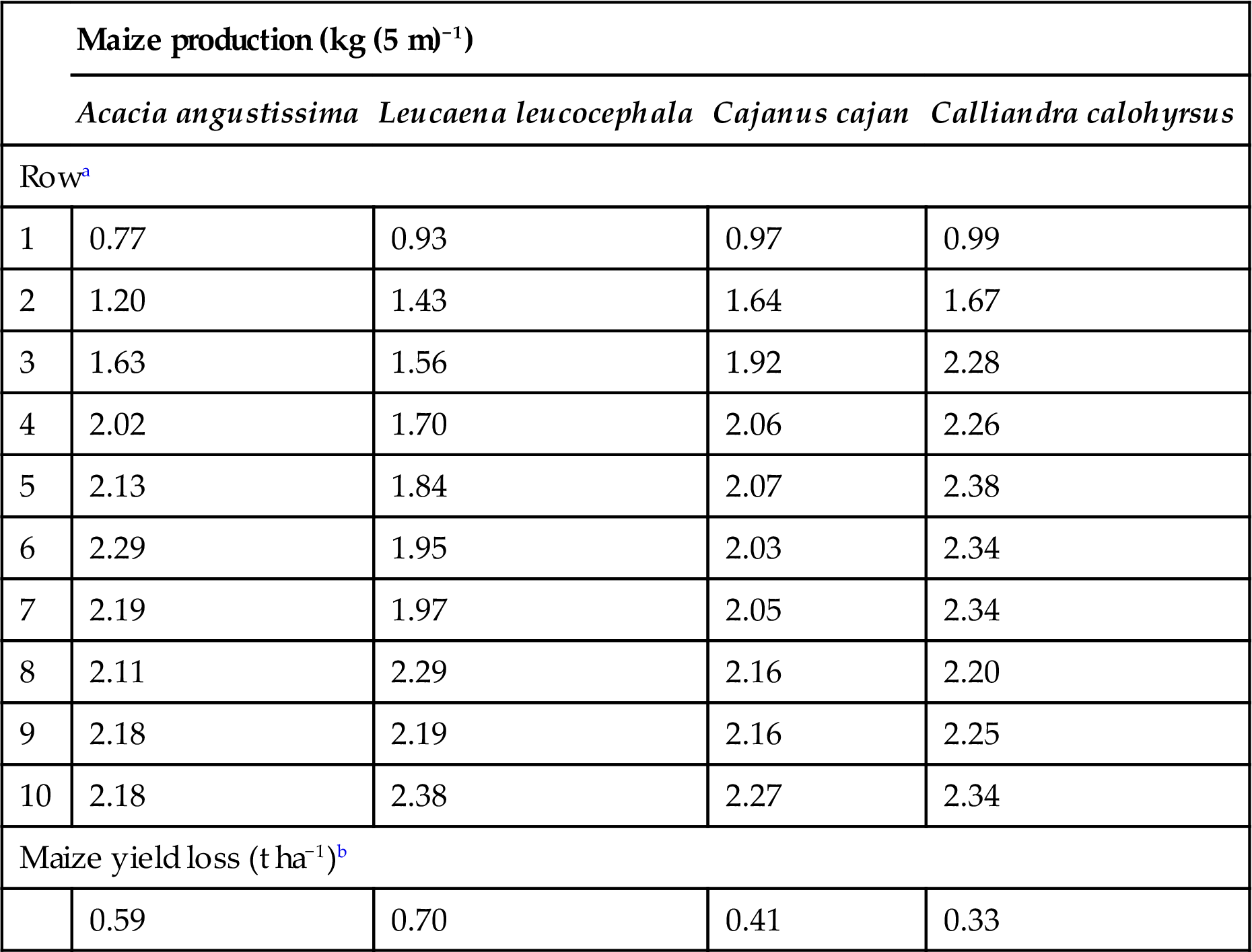
The competition for resources between trees and crops has been discussed in a previous section on hedgerow intercropping, and the same principles hold when trees are planted on conservation structures. For example, studies in Zimbabwe (Dzowela et al., 1994) are evaluating the potential of leguminous hedgerows planted on old earth contour bunds to supply dry season fodder for draft oxen. In addition, the impact of these hedges on the performance of maize grown adjacent to these bunds is being evaluated. The results have shown that such bunds (assuming 500 m bund per farm) do have the potential to provide sufficient fodder for the needs of a 300-kg draft animal, but at the cost of lost maize yield due, in this case, to belowground competition for water (Fig. 4.14). There were differences in the competitive effects of the species evaluated and in the case of leucaena this extended for up to five rows (4.5 m) away from the bund (Table 4.7). In another study in Uganda, Okorio et al. (1994) demonstrated similar competitive effects of linear contour plantings of a range of trees being evaluated for pole production. In this case, they demonstrated that both above- and belowground competition for resources were important and for the most competitive species (Maesopsis eminii) this effect extended for more than 3 m either side of the linear planting. It therefore appears that on steeply sloping land, where conservation structures are closely spaced, the competition between trees and crops will be more important than on more gently sloping land where earth bunds may be spaced up to 20 m apart, as is typical, for instance, in many small-scale farms in Zimbabwe.

Table 4.7
Maize production adjacent to fodder bunds planted with different hedgerow species, Dombashawa, Zimbabwe, 1994.
| Maize production (kg (5 m)−1) | ||||
| Acacia angustissima | Leucaena leucocephala | Cajanus cajan | Calliandra calohyrsus | |
| Rowa | ||||
| 1 | 0.77 | 0.93 | 0.97 | 0.99 |
| 2 | 1.20 | 1.43 | 1.64 | 1.67 |
| 3 | 1.63 | 1.56 | 1.92 | 2.28 |
| 4 | 2.02 | 1.70 | 2.06 | 2.26 |
| 5 | 2.13 | 1.84 | 2.07 | 2.38 |
| 6 | 2.29 | 1.95 | 2.03 | 2.34 |
| 7 | 2.19 | 1.97 | 2.05 | 2.34 |
| 8 | 2.11 | 2.29 | 2.16 | 2.20 |
| 9 | 2.18 | 2.19 | 2.16 | 2.25 |
| 10 | 2.18 | 2.38 | 2.27 | 2.34 |
| Maize yield loss (t ha−1)b | ||||
| 0.59 | 0.70 | 0.41 | 0.33 | |
SED for maize yield loss=0.138.
aRow 1 adjacent to contour bund, Row 10 furthest away.
bMaize yield loss was calculated utilizing the mean yields of Rows 7, 8, and 9 as the control, and assuming 500 m bund length ha−1.
Source: From Dzowela, B.H., Mafongoya, P.L. Hove, L., 1994. SADC-ICRAF Agroforestry project, Zimbabwe. 1994 Progress Report. ICRAF, Nairobi.
In conclusion, high-value trees and shrubs do have an important role to play in both stabilizing conservation structures and increasing their productivity and diversity of output. However, this will almost always be at some cost in terms of reduced productivity of associated crops. Research priorities are those associated with the determination of local community needs and opportunities, and the identification of appropriate species and their management, rather than the impact of trees and shrubs on soil erosion control per se.
Multistrata Systems
Multistrata agroforestry systems have evolved primarily to generate income and provide and diversify basic family food needs. Their function and productivity in these roles is discussed in a subsequent section (see section on high-value trees). However, because of their structural resemblance to secondary forests, they exhibit a profound capacity to prevent soil erosion (Kiepe and Rao, 1994).
Little research has been undertaken to investigate the detailed mechanisms involved but from available evidence it is clear that it is the understory and litter which play the primary role in soil conservation, and that the upper-story canopy has little, if any, beneficial effect. Raindrops reach 95% of their terminal velocity in 8 m free fall from an upper-story canopy, frequently with increased drop size and greater soil erosivity (Soemwarto, 1987). The relative importance of the different layers in multistrata systems was illustrated by a study of erosion under a 5-year-old Acacia auriculiformis stand in Java. The removal of the canopy and understory, but the retention of the litter alone, reduced soil erosion by 95% compared with that experienced from bare soil. The retention of the canopy alone increased the erosive power of raindrops by 24% (Wiersum, 1985). Other evidence for the importance of litter was provided in a comprehensive review by Wiersum (1984). Erosion losses reported in undisturbed forest plantings in the humid and subhumid tropics range from 0.02 to 6.20 t ha−1 a−1 compared with 5.92 to 104.8 t ha−1 a−1 where the understory was burned and the litter removed. These values can be compared with erosion in multistory gardens, which ranged from 0.01 to 0.14 t ha−1 a−1. Indirect evidence was also recently obtained from Uganda where C. calothyrsus and L. leucocephala were established at high populations (1×0.5 m) on 60% sloping land. After 3 years a dense canopy of about 3 m in height and a thick layer of litter had formed. Observations indicated that, even on such steep slopes, soil erosion had been completely eliminated (ICRAF, 1994, pp. 101–108).
Multipurpose Trees and Shrubs as Fodder Supplements
Feed Quality
Shortages of high-quality dry season fodder supply have been widely recognized as a constraint to ruminant production in the tropics of Africa, both for small and large ruminants (Fig. 4.4). Crop residues are utilized as fodder and seldom returned to the soil, and animal manure is often used as a fuel. Such practices exacerbate the negative nutrient budgets of crop production fields and contribute substantially to the steady decline in soil organic matter (Fig. 4.5). In addition, natural pastures and crop residues are of low feed quality, and animal condition and performance almost always declines during the dry season. Already a serious constraint to many small-scale farmers, this situation is projected to worsen dramatically in the future as population pressure continues to build.
A recent extensive review of animal agriculture in sub-Saharan Africa concluded that dwindling feed resources will be the greatest constraint to the productivity of an expanding livestock population (Winrock International, 1992). It is forecast that unless feed production trends improve dramatically, the current 140,000 t meat imported annually into the region (costing US$200×106) will rise from the present level of 3% of the total requirements to about 35% of the region’s needs, or 7×106 t meat by the year 2025. Similarly, milk product imports are predicted to rise from the current level of 11% of total requirements (costing US$500×106 annually) to about 25% by the year 2025. The combined cost of these projected imports of meat and milk products is estimated at a staggering US$16×109 annually, beyond the realms of economic feasibility.
Given the inevitable decline in availability of already poor quality grazing areas and increased pressure on-farm feed resources, a major research and development initiative is required to improve the supply of quality feed at the farm level. Without such an initiative, the projected pressure on the natural resource base to provide livestock feed will result in further widespread land degradation.
However, most small-scale farmers cannot afford or do not have access to concentrated feed which, even when available, can be of variable quality. Legume forage, however, which has a similar effect to commercial concentrates on feed utilization and animal performance, has proved to be adoptable on smallholder farms. Leguminous trees and shrubs have advantages over herbaceous legumes inasmuch as they are persistent, produce more edible dry matter and usually retain their leaves during the crucial dry season period; indeed, they can be deliberately managed to optimize leaf dry matter production during this period. However, many browse species contain antinutritive factors, such as saponins, alkaloids, and tannins, that can significantly reduce their nutritive value and may affect palatability. Condensed tannins have a significant effect on fermentability in the rumen, but have little effect on palatability. For example, C. calothyrsus contains a high level of condensed tannin and is highly palatable when fresh, but has a low level of fermentability in the rumen and low digestibility in the postrumen intestinal tract. Drying and wilting reduces total phenolics and condensed tannin levels, but depresses palatability, degradability and digestibility (Palmer and Schink, 1992; Ahn et al., 1989). G. sepium, on the other hand, which has a low level of tannin, is more palatable after wilting to animals unaccustomed to the feed, but both fresh and wilted material is highly palatable to animals that are used to the feed (Ash, 1989; Smith et al., 1994).
There is also wide variation in the site and extent of breakdown of protein from browse species (Table 4.8). For example, drying increases the rumen degradability of protein and the proportion of undegradable protein that is catabolized in the small intestine in albizzia, erythrina and leucaena, but decreases it in gliricidia and calliandra. Time spent in the rumen is affected by the level of food intake and the rate of breakdown of particles in the rumen.
Table 4.8
Concentrations of protein (CP) and coefficients of rumen degraded protein (RDP), intestinally digested protein (IDP) and excreted dietary protein (undegraded and undigested, ExDP) for various browse species, as affected by forage preparation, forage regrowth period, and length of time the material remains in the rumen.
| Proportions of | ||||
| CP | RDP | IDP | ExDP | |
| (g (kg DM)−1) | ||||
| Forage preparationa | ||||
| Erythrina verigata | ||||
| Fresh | 253 | 0.64 | 0.25 | 0.11 |
| Sun-dried | 253 | 0.74 | 0.20 | 0.05 |
| Gliricidia sepium | ||||
| Fresh | 261 | 0.69 | 0.25 | 0.06 |
| Sun-dried | 261 | 0.70 | 0.19 | 0.11 |
| Leucaena leucocephala | ||||
| Fresh | 246 | 0.39 | 0.37 | 0.24 |
| Sun-dried | 246 | 0.48 | 0.34 | 0.18 |
| Calliandra calothyrsus | ||||
| Fresh | 256 | 0.71 | na | na |
| Dried 25°C | 256 | 0.39 | na | na |
| Dried 65°C | 256 | 0.32 | na | na |
| Forage regrowth time before harvestingb | ||||
| Callianda calothyrsus | ||||
| 12 weeks | 282 | 0.59 | 0.32 | 0.09 |
| 24 weeks | 253 | 0.58 | 0.26 | 0.16 |
| 48 weeks | 231 | 0.51 | 0.23 | 0.26 |
| Forage time in the rumenc | ||||
| Sesbania sesban | ||||
| 24 h | 267 | 0.77 | 0.10 | 0.13 |
| 48 h | 267 | 0.84 | 0.06 | 0.10 |
| Leucaena leucocephala | ||||
| 24 h | 257 | 0.44 | 0.32 | 0.24 |
| 48 h | 257 | 0.57 | 0.21 | 0.22 |
| Calliandra calothyrsus | ||||
| 24 h | 217 | 0.24 | 0.31 | 0.45 |
| 48 h | 217 | 0.31 | 0.25 | 0.44 |

aPerera et al. (1992), Palmer and Minson (unpublished).
bKaitho et al. (1993).
cKamatali et al. (1992).
For all browse species studied, an increase in the time spent in the rumen raises the level of rumen degradability and decreases the level of subsequent catabolism in the small intestine. The proportion of dietary nitrogen excreted in dung or lost in urine from livestock on a forage diet will depend upon the site of catabolism in the digestive tract (Reynolds and de Leeuw, 1994). It is not biologically efficient to include more rumen-degradable protein in a diet than can be trapped by the rumen microbes. This is particularly true in farming systems where manure is essential for soil fertility maintenance (e.g., those in the highlands of Rwanda) since nitrogen in dung is more easily managed than nitrogen in urine. Urine cannot be easily handled by farmers, and there is a high loss of nitrogen from urine through volatilization and leaching. It would be more efficient (in terms of nitrogen utilization) to compost surplus high-nitrogen vegetation than to pass it through animals. However, as previously indicated, the economics of green manure versus fodder supplementation need to be carefully assessed.
In a study of three browse species using cattle, sheep and goats, Smith and colleagues (1994) have found highly significant browse x animal species interaction for the rate and extent of degradation of the slowly degradable dry matter fraction in browse. Since there is no difference in the capacity of sheep and goats to digest most conventional diets (grasses, crop residues, concentrates), except when dietary protein levels are very low (Tolkamp and Brouwer, 1993), this suggests specific adaptation by different animal species to browse species. Care must therefore be taken in extrapolating nutritive values for browse species from one animal species to another.
Effects on Animal Performance
The addition of leguminous browse species to a basal grass ration can lead to a marked improvement in animal performance. High-quality browse can be grazed in situ (e.g., as practiced for beef cattle in Australia) or it can be offered as a cut-and-carry supplement to stall-fed animals (e.g., dairy cattle on smallholder farms in Kenya). Cattle growth rates, e.g., improved from −20 g d−1 on natural pasture in Indonesia to +540 g d−1 when leucaena comprised 40% of the ration (Wahyuni et al., 1982). Milk production of dairy cattle on smallholder dairy farms in Kenya increased by 0.5 kg milk per kg leucaena added to a diet of Pennisetum purpureum (Fig. 4.15). During early lactation the response in milk production is high, particularly in the dry season when grass quality is poor (Fig. 4.16), but in mid to late lactation the milk response is more muted as a greater proportion of additional nutrients are directed toward body tissue (Muinga et al. 1995). In Embu, in Kenya, the replacement value of C. calothyrsus as a fodder supplement has been found to be in the region of 3 kg fresh material (=1 kg dry matter), equivalent to 1 kg of dairy meal with 16% crude protein for grade Friesian and Ayrshire milking cattle (R. T. Paterson, unpublished data). However, for local Ankole cattle in Rwanda with low milk productivity, it has been shown that there is little economic gain to be had through the use of high-quality tree fodder as a supplement (Niang et al., 1996). The use of tree fodder for increased milk production must be judged according to the productivity and hence nutritional needs of the animals involved. An improvement in body condition should increase reproductive performance by raising conception rates and reducing calving intervals, although direct evidence from long-term trials with legume forage is lacking.

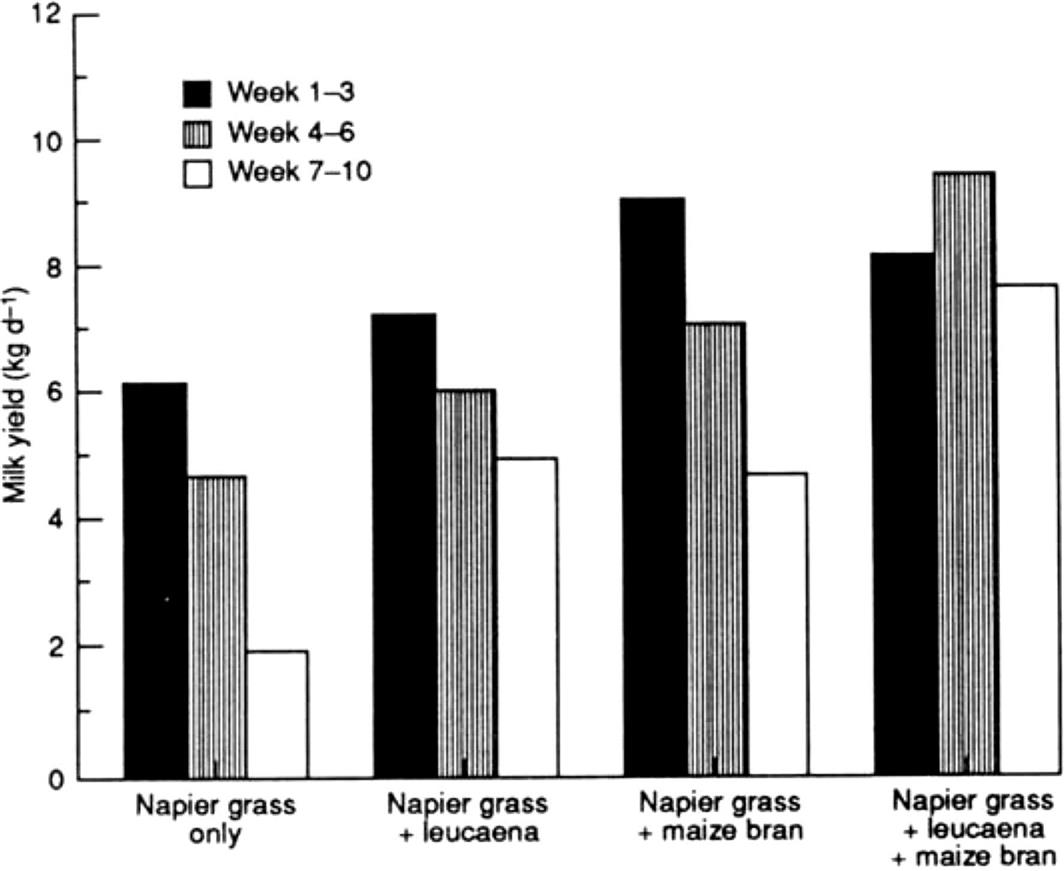
Small ruminants also benefit from the inclusion of leguminous browse in their diet, with faster growth rates and improved reproductive performance. The most significant change following supplementation of pregnant and lactating dams is in the survival rates of their offspring, which rises from 45% on a grass-only diet to over 95% when browse comprises 40% of the ration (Reynolds and Jabbar, 1994). Significant health and nutrition interactions were observed, increasing survival rates, when browse supplements were offered to sheep infected with trypanosomiasis (Reynolds and Ekurukwe, 1988). This is a nutritional effect rather than a direct consequence of browse per se but for many farmers browse is the most available option for improving the quality of the ration in the dry season. Several studies have confirmed that the supplementation of native browse with high-quality fodder also enhances the dry matter intake and live-weight gains of young goats and sheep. In Rwanda, Mimosa scabrella fed to young goats as a supplement to a basal diet of Setaria splendida increased their live-weight gain from 31 to 50 g d−1 (ICRAF, 1994, pp. 98–101). Similar results have been reported from Zimbabwe when Acacia angustissima was used as a fodder supplement for local goats browsing native dry season pastures. Live-weight gains were dramatically increased from −20 to +12 g d−1 during the 50-day period of the trial (Dzowela et al., 1994).
Fodder Production Systems
Three issues are of particular concern in the development of improved fodder production systems for small-scale farmers. The first is the identification of the available niches on farms for the introduction of fodder trees. Many exist and the choices farmers make will determine both the spatial arrangement and, to some extent, the management of the trees. The second issue is that of tree management to optimize fodder production at critical times of the year, which in most instances occurs during the dry season. The third is the choice of species.
In many instances, opportunities exist for linear planting of fodder trees on farms as internal hedges, in combination with upper-story trees as boundary plantings, along terrace risers on sloping land or along permanent contour bunds. Fodder trees can also be planted in blocks either as pure stands or interplanted in existing grass fodder banks (O’Neill, 1993). Furthermore, trees can be planted in cropland either as scattered trees (ICRAF, 1992b, pp. 83–84) or in a hedgerow intercropping arrangement (Reynolds and Jabbar, 1994).
The management of trees to optimize fodder production will be specific to species, spatial arrangement, environment, and interaction with associated trees or shrubs. Where trees are planted in linear arrangements, a within-row tree spacing of between 0.25 and 0.5 m is recommended and experience suggests that pruning fodder trees to a height of between 60 and 100 cm is the most appropriate with regard to ease of labor, quantity of fodder produced, and competition with nearby crops (Niang et al., 1996). At Embu, in Kenya, in a bimodal rainfall regime, when L. leucocephala was pruned at intervals of about 3 months to a consistent height of 100 cm, initial leaf yields were highest when side branches were pruned back to the main stem. Annual cumulative yields were increased, however, by cutting the side branches at about 20 cm from the stem. The yields could be further increased by hand-stripping the remaining leaves after the pruning but the extra labor this involved may not be justified by the small increase in dry matter production (ICRAF, 1994, pp. 109–111). Production is also enhanced if trees are allowed to develop strong root systems before pruning.
Studies have shown that fine-root die-back occurs when fodder trees are pruned (e.g., Reynolds and Jabbar, 1994), so that belowground root regrowth is needed in addition to aboveground fodder production. Pruning too frequently over a prolonged period will reduce fodder production.
In the highlands of East Africa, the contour planting of fodder grasses (P. purpureum below 2000 m altitude and Trypsacum laxum above 2000 m) has been widely adopted by farmers as a dual-purpose erosion control and fodder production system (Fig. 4.17). High-quality fodder-producing trees can easily be established at the same time as the grass strips or within established lines. Research has shown that the inclusion of such trees in combination with grass strips dramatically increases the production of digestible protein without affecting total dry matter production (Niang et al., 1996; ICRAF, 1995). For example, in Burundi the productivity of two contour-planted rows of trypsacum grass was compared with a single row of trypsacum combined either with a row of C. calothyrsus or Leucaena diversifolia (Table 4.9).

Table 4.9
Fodder production from different combinations of grasses and shrubs in Burundi.
| Mean annual production 1991–93 (kg DM m−1) | Mean annual production of digestible protein (g m−1) | |
| Two rows trypsacum | 2.22 | 89 |
| One row trypsacum + one row calliandra | 2.06 | 179 |
| One row trypsacum + one row leucaena | 1.83 | 119 |
Source: From ICRAF (1995).
From these results it can be calculated that a Burundian farmer would only need to plant 60 m of a calliandra/trypsacum hedge to meet fully the annual maintenance and digestible protein requirements of a local 20-kg goat. Such an approach is being widely adopted by milk producers near Embu in Kenya where farmers are planting calliandra in established contour plantings of pennisetum and are reporting significant increases in milk yield. Other research in Zimbabwe has demonstrated that A. angustissima, grown on existing 2-m-wide contour bunds which average 500 m bund length per hectare of cropped land, can provide the crucial supplementation needs of draft oxen during the dry season (Dzowela et al., 1994). Fodder production on these bunds is now being evaluated with local farmers for a range of adapted fodder species (Fig. 4.14).
Fodder trees can also be grown as linear hedgerows in crop fields in an identical arrangement to that described for hedgerow intercropping. Where such hedgerows are planted on the contour for erosion control, hedgerow prunings can be most economically utilized for fodder under mildly erosive conditions (see previous section on barrier hedges) and this is an attractive option for farmers. However, on flat land such an approach has little to offer. Removal of prunings for fodder instead of their use as a green manure will inevitably result in a reduction in associated crop yield. This could largely be avoided if the same number of trees were planted as a block in part of the field, since this arrangement results in a reduced zone of tree/crop interface. In such block plantings, where trees are commonly spaced at approximately 0.5×1.0 m, fodder yields range from 0.5 to 1.0 kg tree” a−1 in the humid and subhumid tropics. The recommended level of fodder supplementation for a grade dairy cow is about 2 kg dry matter d−1 or 700 kg a−1. Between 700 and 1400 trees are therefore required per lactating animal, or a land area of between 0.035 and 0.07 ha only.
In many areas, particularly those with long dry seasons, farmers need to adopt a tree pruning strategy that maximizes fodder production at periods of greatest shortage. Under such conditions moisture stress and leaf senescence can lead to the loss of fodder material unless cutting times are chosen judiciously. If, e.g., the aim is to maximize the yield of fodder at the height of the dry season in August, as in Burundi, research has shown that the final wet-season cut should be made 6 months earlier in February (Akyeampong and Muzinga, 1994). Earlier cutting resulted in increased leaf senescence while later cutting gave a suboptimal period for recovery before the onset of the dry season.
Cultivation of fodder trees on grassland is only feasible where grazing access can be controlled. For example, over 200 km2 of natural pasture on commercial farms in Queensland has been strengthened with leucaena for use by beef or dairy herds (Wildin, 1994). On communal land or where livestock graze freely after crops have been harvested, young fodder trees can be killed by free-roaming animals. In some parts of Africa, however, farmers collectively impose control over special areas set aside for dry season grazing. An example is the Ngitiri system practiced by the Sukuma peoples in the Shinyanga district of Tanzania (Otsyina and Asenga, 1993). Collectively or individually owned areas are protected during the rainy season and are then grazed with careful regulation during the dry season by draft oxen. However, farmers recognize the generally poor quality of the dry season grazing and have expressed considerable interest in establishing adapted and productive browse species in their Ngitiri. They have indicated that linear plantings of such species, to subdivide Ngitiri areas into paddocks, would further assist in dry season grazing control. L. leucocephala, G. sepium, A. angustissima, C. calothyrsus, and Atelia herbertsmithii are currently being evaluated together with farmers.
The choice of species will depend not only on their feed quality, as discussed earlier, but also on their specific adaptation and their capacity to remain productive under continuous pruning. S. sesban, e.g., is a widely adapted and fast-growing species with high-quality fodder. However, experience has shown that its regrowth after pruning is poor and early death can result (Niang et al., 1996).
In general, it has been found that adapted exotic material shows greater vigor and fodder productivity than indigenous species. In many parts of sub-Saharan Africa with relatively neutral soils, L. leucocephala was a most productive fodder species and was becoming popular with farmers until the arrival of the leucaena psyllid (Heteropsylla cubana) into the region. This pest first appeared on the African mainland just north of Mombasa (Kenya) in August 1992 (Reynolds and Bimbuzi, 1993). It has recently appeared in Zimbabwe and its presence in neighboring countries would suggest that it will become endemic in the whole region before long. It remains to be seen if natural or, possibly, introduced predators will be able to control it in the long run. In Asia, the impact of the psyllid was much reduced 4–6 years after its initial invasion and yields of leucaena biomass are now almost equal to preinfestation levels (van den Beldt and Napompeth, 1992). Other species such as L. diversifolia, Leucocasia esculenta, and Lambula pallida, together with a number of composites and interspecific hybrids, have shown some degree of tolerance to the psyllid (Dzowela et al., 1994; Otsyina et al., 1994a), but work with them is of recent inception and more local experience will have to be gained before these species and lines can be recommended to farmers. Of other exotic leguminous species tested, those with widest general potential across the region would appear to be C. calothyrsus where the annual rainfall exceeds 1100 mm and G. sepium where the annual rainfall is less than 1100 mm but above 700 mm. On acid soils in Zimbabwe the cold tolerance of A. angustissima makes it a very promising species (Dzowela, 1994) while at altitudes above 2300 m on extremely acid soils (pH below 4) in Rwanda M. scabrella has been shown to outperform most other species (ICRAF, 1994, pp. 98–101).
Selection work with indigenous fodder trees is not as well advanced as with exotic species. Trees which are valued by farmers as sources of fodder include both legumes and nonleguminous species. In the bimodal rainfall highlands of Kenya, farmers value the local species Trema orientalis and Sapium elipticum while the naturalized species Morus alba, which is commonly recognized as a source of feed for silkworms, also shows potential for the feeding of ruminant livestock (Thijssen et al., 1993). In eastern Zambia the local species Zizyphus abyssinica and Diplorynchus condylocarpon have been shown to be useful supplements for goats fed on a basal diet of poor quality roughage (Phiri et al., 1992) while in Tanzania Margaritaria discoides is showing similar promise for goats (Otsyina et al., 1994b).
In conclusion, considerable information is available on the potential of trees to provide high-quality fodder, their adaptation to specific environmental conditions, the range of farm niches where they can be planted and appropriate management regimes to optimize their production and impact. In spite of such information, widespread adoption is currently limited to peri-urban milk producers where market forces provide a clear incentive and farmers are already aware of the impact of feed supplementation on milk yields. Where oxen are used for draft power and provide a source of income for farmers (as in Zambia and Tanzania), farmers are interested in feed supplementation during the dry season but widespread adoption has yet to occur.
In the future, as urban populations increase and natural grazing areas are unable to support expanding herds of beef cattle, it is probable that peri-urban beef industries will develop, as is already being observed for poultry and egg production. The rate at which such systems develop and their economic viability will depend upon a reliable and cheap source of high-quality feed supplement. Fodder trees will have an important potential role (Winrock International, 1992). However, research will need to address the issue of the sustainability of such tree-based fodder production systems since they will result in high rates of nutrient export (Shepherd et al., 1994). Efficient nutrient recycling of animal waste is likely to be important in this respect.
High-Value Trees for Income Generation
In the last few decades, since emerging from being just a traditional form of land-use, agroforestry has been viewed as a panacea for the achievement of sustainability and land amelioration. More recently, however, agroforestry has reemerged as a means of producing traditionally important and valuable forest products such as timber, fruits, medicines, and extractives on farmland. In this way agroforestry can now be viewed as an intervention point to break the downward spiral of land degradation and rural poverty (Fig. 4.5). The production of forest products can be used either to generate cash with which to buy fertilizers to increase the yields of staple crops or as a profit-motivating incentive to promote the establishment of more trees on-farm to ameliorate soil depletion and land degradation. Thus the vision now is of agroforestry as an integrated land-use policy that combines increases in productivity and income generation with environmental rehabilitation and the diversification of agroecosystems. Such a vision can be fitted to the range of situations found in the major ecoregions of the tropics. The realization of this vision, however, is going to depend on three factors in particular. The first is an appreciation by the international community and donors of the importance of high-value indigenous species in the lives and welfare of local people, as well as an incentive (or the removal of disincentives) for local people to plant trees on their farms. The second is domestication of commercially important indigenous tree species producing forest products, and the third is the development of a processing and marketing infrastructure.
Naturally Regenerating Trees in Farmland
In Africa, there are still relatively few examples where trees are being deliberately established on farms to generate income. While examples of farmers growing trees for this purpose are known to the authors, there is very little published evidence of the scale of such activity in different areas of Africa. It is, however, quite common for farmers to protect and retain naturally regenerating trees on farmland, both for the production of a regular income from fruits and as a capital investment in the form of quality timber which can be liquidated to pay school fees or wedding/medical/funeral expenses.
In the Sahelian parklands, large trees of Butyrospermum paradoxum and Parkia biglobosa are grown as scattered trees in the fields, and the fruits of these trees are harvested and marketed. Despite a reduction in the production of crops under the trees, the overall returns per hectare are increased by 6500 FCFA ha−1 (250 FCFA=US$1) with one B. paradoxum and one P. biglobosa per hectare and by 13,600 FCFA ha−1 if the number of B. paradoxum is increased to eight (Bonkoungou, 1995). Although these economic benefits are not large, they do represent an important cash income to relatively poor farmers.
In southern Africa the indigenous fruits of the miombo woodlands are also important and are beginning to contribute to commercial trade. Data are scarce on the value of this trade but, e.g., the US$103 made by a marketeer in Zambia in September 1993 from the sale of Ziziphus mauritiana, one of the locally marketed fruits (Kwesiga et al., 1994), is significant when the average annual per capita income for Zambia is only $272.
In the humid forests of West Africa there are also numerous species the products of which are traded (Fig. 4.18). For example, cola nuts and the kernels of the bush mango (dika nut) are sold on a substantial regional scale. Again the economic value of this trade is not well documented. The cola nut trade in 1981 was reported to be 22,500 t (Nkongmeneck, 1985) of which 20,400 t entered commerce, mostly being traded to the north where it is the only stimulant allowed to Muslims. Data on bush mango kernels from Cameroon, however, show that the price per kilogram fluctuates seasonally from a low in August (535 FCFA) to a high in February (1670 FCFA), with farmers receiving 50–72% of these urban prices in July–August but only 18–39% in August–October (Ndoye and Tchamou, unpublished). The trade in some other products, such as the chewing sticks from Garcinia species, has been quantified. In this case the street value from Kumasi market in Ghana was US$9×106 a−1 (Falconer, 1992). In all these examples the marketing has developed to meet local and regional demands and is based on traditional practice.

Another example, and one involved in international trade, is “pygeum” (Prunus africana) which grows in discontinuous populations throughout sub-Saharan Africa in the montane areas above 1200 m and where rainfall exceeds 1000 mm. The bark of this tree is harvested for pharmaceutical products used in the treatment of benign prostatic hyperplasia and prostate gland hypertrophy, ailments suffered by 60% of men in Europe and the United States (Fig. 4.19). The market value of the commercial product has been estimated at US$150×106 a−1 (Cunningham and Mbenkum, 1993). The overexploitation of this tree’s bark is seriously threatening the resources of the species, with implications for the future source of the pharmaceutical product, as it is apparently unlikely that this drug will be synthesized, as well as for the montane ecosystems of sub-Saharan Africa. This tree, which also produces good timber, is beginning to be grown by farmers in Cameroon and has potential for agroforestry.

Planted Trees in Farmland
The best example of farmers deliberately growing indigenous trees for cash generation is in Southeast Asia, not in Africa. Some Asian examples are included in this review to illustrate the potential of the technologies, from which we believe Africa can learn.
In southwest Sumatra, near Krui, farmers have for more than a century been planting damar trees (Shorea javanica) for resin production, in mixtures with a range of indigenous fruit tree species. The area now exceeds 10,000 ha and 65–80% of the households are involved in damar production (Fig. 4.20). In 1994, the production was expected to reach 10,000 t (Dupain, 1994) at a value of US$0.3–0.4 kg−1. These trees are planted among rice, coffee, and other crops, so that at all stages of their growth the farm is productive—food crops in years 1–3, coffee and bananas in years 3–8, fruits and fuelwood in years 2–20, and resins, fruits and timber from year 20 onwards (De Foresta and Michon, 1994). In the case of damar, the resins are utilized by industries in Indonesia or exported worldwide. In 1984, the export market represented one-third of the harvested volume, and trade increased from 250 to 400 t a−1 between 1972 and 1983 (Michon and de Foresta, 1995). Most of this trade (80%) is met by the damar agroforests. The economic value of the damar trade and its associated activities is of major significance to the villages around Krui. In 1993, the profits from damar production were US$7.25×106 (US$3.25×106 from sales, US$2.65×106 from added value and US$1.35×106 from wages). To this should be added US$271,000 profit made by Krui traders (Michon and de Foresta, 1995). This analysis excludes the locally consumed products from these agroforests such as fruits, vegetables, spices, fuelwood, timber, palm thatching, rattan, bamboo, and fibers.

Systems similar to the damar agroforest are also practiced in Sumatra with rubber and cinnamon. The “jungle rubber” agroforests covered more than 2.5×106 ha while there were 42,600 ha cinnamon in 1989 (Aumeeruddy, 1994). These damar, cinnamon and jungle rubber multistrata agroforests are probably the ultimate example of an alternative to slash-and-burn agriculture, being highly productive, biologically diverse and very similar in structure to natural tropical high forest. In short, they are the nearest thing to a sustainable manmade ecosystem.
Probably the best example of the economic value of planted trees in an agroforestry system on a small farm in Africa is the cost/benefit analysis of 18-year-old G. robusta trees in boundary plantings in Embu, Kenya (Tyndall, 1996). Here at 1500 m altitude, with a bimodal rainfall pattern of 1300 mm a−1 and on deep nitisol soils, G. robusta is grown on farms producing maize, beans, coffee and milk and averaging 1.5 ha in area. Boundary plantings resulted in 85% of households being self-sufficient for fuelwood (Fig. 4.21). The trees are grown mainly for firewood and timber in a single row along the edge of a field at a spacing of 2.7 m. This cost/benefit study examined the inputs and outputs over an 18-year period and discounted these at 20% in a 1-ha plot of maize in which 37 trees were grown down one side of the plot. This was compared with a 1-ha plot without trees. In the 13 m adjacent to this row of trees the maize suffered from competition, resulting in an average loss of 5% over the whole hectare. The G. robusta plots were found to have added benefits that totaled US$87 ha−1, while the added costs totaled US$79 (Table 4.10). Thus the discounted net benefit was US$8 ha−1 (10% greater than added costs). This, like the indigenous fruits example, highlights the fact that in purely economic terms the benefits of even high-value trees only just exceed the associated costs and yield losses. Despite this, farmers are planting grevillea widely. Indeed, although higher elevation divisions of Embu (Manyata and Runyenjes) have some of the highest human population densities in Africa, they also have rapidly growing tree populations as farmers are adopting agroforestry. The unquantified benefits probably include reduced risk, reduced pressures on neighboring forests, erosion control on sloping land, shade and amenity, freedom from boundary disputes and enhanced climate effects (e.g., a windbreak effect).

Table 4.10
Cost/benefit analysis ($ ha−1) at net present value of 18-year-old boundary plantings of Grevillea robusta.
| Added benefits | Added costs | Net discounted benefit | ||||||
| Fuelwood | Timber | Saved inputs | Total | Lost maize | Potential yield lost | Nursery cost | Total | |
| 47 | 29 | 11 | 87 | 47 | 28 | 4 | 79 | 8 |

From Tyndall, B., 1996. The Anatomy of Innovative Adoption: The Case of Successful Agroforestry in East Africa. Ph.D. Thesis, Colorado State University, 212pp.
Data on the yields of crops planted adjacent to rows of tree crops are relatively scarce but in one study in Uganda 17 upper-story tree species were compared for their variability in competition with associated crops (Okorio et al., 1994). The relatively fast-growing trees with broad and dense canopies (M. eminii and Cordia abyssinica) or tall canopies (M. lutea and Melia azedarach) appear to be more competitive than those that are slow growing (A. acuminata) or those with narrow and lax canopies, suggesting that shading is the major source of competition. Relatively little is known, however, about the belowground competition, although species like M. eminii, in contrast to A. acuminata, have large amounts of fine feeder roots in the top 30 cm of the soil. In addition to this species variation in belowground architecture, it is likely that there is variation within-species. The identification and development of methods to utilize this variation is an area for future research. In the Ugandan study A. acuminata was the only species that had an apparent positive effect on crop performance. This was attributed to nitrogen fixation by its Frankia symbionts, since soils sampled from under this species were significantly richer in nitrogen, soil organic matter, calcium, and potassium, but conclusive evidence of the mechanism involved is lacking.
Home Gardens
The intimate association of trees, crops, and animals in home gardens makes them a good example of an agroforestry system. The outputs from home gardens are diverse and production provides farmers with extra income. Unfortunately, in Africa home gardens are not well developed although they exist in many areas (Fig. 4.22). In Nigeria, e.g., they are important in areas with high population densities (1000 people km−2). In such areas up to 29% of the cultivated area can be in compound gardens (Okafor and Fernandes, 1987) and these produce 59% of the crop output (Watson, 1990). In monetary terms the output of these gardens is 5–10 times greater than that of crop fields, with returns to labor 4–8 times greater. Despite this, most of the examples quoted in this review are again from Southeast Asia or from Latin America. Many of the biophysical interactions taking place in home gardens are complex, difficult to research and poorly understood. Despite this, it is probably fair to claim that home gardens are among the most sustainable of land-use systems and that they have the following characteristics (Torquebiau, 1992).

Home gardens are generally accepted as having a high biological diversity. For example, in Java 270 plant species have been recorded in the 41 home gardens of one village while 338 plant species were reported in one area of Mexico. In addition, it is common to find local varieties of tree crops established in home gardens, e.g., in Micronesia 21–37 varieties of each of three species (coconuts, breadfruit, and bananas) have been recorded. In Mexico 15% of the woody plants were locally wild species. A correlation has been found between the diversity of home gardens in Mexico and the available labor force. These home gardens are also biologically diverse in terms of wildlife. In Java, e.g., out of 121 different bird species identified in home gardens in four villages 15 were endangered species.
From the socioeconomic viewpoint food production from home gardens gives good returns on labor. In Indonesia 7% of people’s time is spent in the home garden but this effort produces 44% and 32% of their total intake of carbohydrates and proteins, respectively. This production can be achieved at low cost. For example, in Java production costs only represent about 10% of the output or 15% of the gross income, while in paddy rice equivalent values are between 30–50% and 56%, respectively. Nutritionally, home gardens are also very important. In Hawaii, e.g., a 35-m2 home garden can provide 100% of the vitamin A and C requirements, 50% of the iron requirements and 18% of the protein requirement of a family of five. Thus, as in Java, the home garden species supply the nutrients in shortest supply from the staple diet of rice. Reported values for total calorie and total protein intakes provided by home gardens are variable, with values as high as 44% and 32%, respectively. Interestingly, during 1975–83 fruit prices rose 10-fold in Indonesia, while the price of rice rose by only 2.4 times, indicating that the market for diverse tree-crop products may be increasing relative to that for staple commodity crops.
Production from home gardens is not, however, limited to food crops. Levels of wood production of 7–9 m ha−1 a−1 have been recorded in Java, which compare well with growth rates obtained in plantation forestry. In Java 51–90% of fuelwood comes from home gardens, while in Bangladesh it has been noted that 70% of the sawlogs and 90% of the harvested fuelwood and bamboo come from the homestead. In addition the diversity of products from home gardens also provides opportunities for the development of cottage industries. This creates jobs and off-farm employment.
In conclusion, home gardens are both highly productive and sustainable. However, it must be noted that this is, at least in part, due to regular inputs of household, animal and human waste which supplement nutrient uptake by deep rooted trees.
Domestication
Extractive harvesting or gathering, retention of natural trees on the farm and the planting of unimproved germplasm are the first three steps of the domestication of a tree species. Accepting the argument that there is an important niche in agroforestry systems for high-value wild species, a strong case can be made for the further domestication of these “Cinderella” species (Leakey and Newton, 1994a,b; Leakey et al., 1994; Newton et al., 1994). The domestication process is multifaceted (Leakey and Newton, 1994c) and will vary depending on the value of the species, the duration of its reproductive cycle and the length of the agroforestry/forestry rotation to which it is suited. In all cases it is important that a domestication strategy be developed which ensures the maintenance of genetic diversity and the continuing introduction of new cultivars, so that the risks of a narrowing genetic base and stagnating commercial production are avoided (Leakey, 1991; Simons et al., 1994). The development of genetic homeostasis between cultivars, furthermore, allows a deployment strategy in which cultivars can fill different niches within a farm to take advantage of differences in optimal environmental conditions and in competitiveness (Foster and Bertolucci, 1994).
In recent years a number of new tree products have appeared on the supermarket shelves. Kiwi fruit, pecan nuts and macadamia nuts are perhaps the best examples. In the case of macadamia nuts domestication arose from the combination of a breeding program in Hawaii and the clonal propagation of cultivars selected for tree habit, yield and kernel recovery, which are grafted onto canker resistant rootstocks (Cannell, 1989). The newest clones are reputed to yield four times as many nuts per tree and to have 10% more first-grade kernels than the earlier clones.
In the tropics a number of indigenous fruit trees are in the early stages of domestication (Okafor and Lamb, 1994; Clement and Villachica, 1994), with the selection of superior individuals for grafting and the collection of germplasm. These approaches need to be linked to studies to maximize harvest index (Cannell, 1989) and to the integration of trees and crops into food crop systems (Watson, 1983). At ICRAF, the domestication of some indigenous fruit trees has started in Malawi and in Cameroon (ICRAF, 1995; Leakey and Maghembe, 1994). In Malawi field trials have demonstrated that, contrary to expectation, the growth of indigenous fruit trees is quite fast and that Azanza garckeana, Annona senegalensis, Bridelia micrantha, Bridelia cathatica, Z. abyssinica, Z. mauritiana, and Ficus vallis-choudea can start to produce flowers and fruits within 2 years (Maghembe, 1994). The indications are that there are substantial differences between trees in fruit size, color, and flavor, and current work is directed toward capturing this variation by vegetative propagation.
A similar approach with indigenous fruits is in progress in Cameroon but in this instance the first step has been to develop and apply guidelines to determine the species preferences of farmers in Ghana, Nigeria, Cameroon, and Gabon in order to prioritize species for domestication on a regional basis (Jaenicke et al., 1995). In both southern and west Africa, these domestication initiatives have also involved workshops for scientists to gather information on the priority species (Maghembe et al., 1995; Boland and Ladipo, unpublished). The result in west Africa has been the choice of Irvingia gabonensis as the first priority and consequently germplasm collections have been made in each country for the establishment of living ex situ germplasm banks in Nigeria and Cameroon (ICRAF, 1995). Coupled with the establishment of these germplasm collections has been the identification by farmers of the best trees in each village. Scions and cuttings from these trees will be used to establish local cultivars that can be rapidly reintroduced into the villages and established for on-farm testing.
In conclusion, while tree products are one of the main factors determining the inclusion of high-value trees on farms, they are probably the least well studied and understood reasons for farmers’ acceptance of agroforestry. This is particularly the case in Africa, but the examples from Asia and Latin America provide hope for the future.
In many instances the demands of growing human populations, coupled with the rates of deforestation, are leading to the exhaustion of the natural resource and an uncertain future. Agroforestry has the potential to reverse these trends, especially if the incentives to plant can be generated by the genetic improvement of the products, so that farmers can increase their returns on investment, and by the expansion of the markets. Unfortunately, both these precursors of change must occur simultaneously and international agricultural research and commerce are not well integrated. However, the market constraints have recently been receiving considerable attention (Vosti and Witcover, 1995) so there is hope that the desired progress can be made. Given the potential of recently modified horticultural techniques applied to the domestication of the trees which traditionally have only been important as sources of extractive reserves, there is now a new impetus for examining the market potential of the most promising commercial species. Ways will have to be found to promote the marketing of species prior to their domestication as the costs of domestication are high. However, since the greatest potential of these species will be in local urban and regional markets where they are already well known, these costs may be very much less than those associated with breaking into an international market. There are therefore many areas for both biophysical and socioeconomic research for each potential domesticated tree, spanning the full range of processes associated with domestication.
Conclusion
To date, research in sub-Saharan Africa has placed emphasis on the development of agroforestry systems which directly address the maintenance of the soil resource base through enhanced soil fertility and soil erosion control, and good progress has been made. Where the development of such systems are built on indigenous concepts such as fallowing, the chances of adoption and impact appear high. We believe, however, that agroforestry solutions to land degradation are always likely to be location-specific in their relevance and acceptability. For this reason it is imperative that research increasingly includes farmers as partners in the research process.
We introduced this review with a brief overview of projected population increases and their probable impact on land-use and land degradation. The inference is clear. Agroforestry systems that provide solutions for today’s land degradation problems will need to evolve in both diversity and intensity if they are to remain relevant and effective for tomorrow’s Africa. The intensification and diversification of agroforestry systems should be equated to an ecological succession and agroforestry should therefore be seen as a dynamic rather than a static form of land-use. In this connection it is interesting to note that in many areas of high population density, particularly in Southeast Asia, the pathway of evolution of land management following forest clearance has nearly gone full circle. Land originally cleared for shifting cultivation and crop production is now increasingly coming under complex manmade agroforests which closely resemble, both in structure and environmental function, the indigenous forests they have replaced. This phenomenon offers great hope for the deforested and degraded areas of the tropics, especially in Africa, but it is clear that the political and policy environment has to be favorable for this type of change to occur—land tenure in particular is a probable requirement. Increased research emphasis needs to be placed on the domestication of high-value indigenous trees and their integration into more sustainable, diverse, and intense land-use systems. Such research is both long-term and complex, but should be embarked upon now if the research community is to be in a position to meet the needs of future generations.
Acknowledgments
The authors acknowledge, with gratitude, all those who contributed to the preparation of this paper, in particular Dr. K. Shepherd, Dr. C. K. Ong and Dr. R. Coe of ICRAF for their review and useful comments on early drafts, Miss M. Mwangi for all her secretarial input and Mr. A. Njenga who provided many of the photographs.