
3.2 Physiology of the human hearing system
This section briefly describes the most important parts of the human auditory system. For more detailed information the reader is referred to dedicated textbooks, e.g. [226].
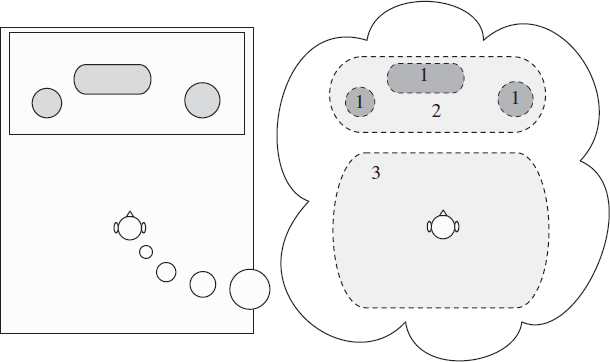
Figure 3.1 Illustration of the perception of the auditory spatial image. Perception of auditory objects (1); extent of the auditory spatial image (2); and listener envelopment (3).
Outer and middle ear
Changes in the acoustical pressure are received by the outer ear, which consists of a visible part (the pinna) and a canal leading to the eardrum (the tympanic membrane). Attached to the eardrum are the middle ear bones (ossicles): the malleus (the outermost), the incus and the stapes. The foot of the stapes is connected to the oval window, which is part of the inner ear. The major function of the middle ear is to ensure the efficient transfer of sound from the air to the fluids of the cochlea. It acts as an impedance-matching device that improves sound transmission.
Inner ear
The cochlea within the inner ear is composed of a bony labyrinth. It is partitioned into three fluid-filled tubes. These tubes are separated by membranes: between the first (scala vestibuli) and second (scala media) there is Reissner's membrane. The scala media is in turn separated from the lower space (scala tympani) by the basilar membrane.
The so-called organ of Corti spirals on the basilar membrane. Two types of sensory cells can be differentiated by their position within the organ of Corti: inner hair cells (IHCs) and outer hair cells (OHCs). The inner hair cells make up a single row of receptors, while the outer hair cells are much greater in number and are often organized in three rows.
The main task of the cochlea is to analyze sound in terms of its intensity, timing and frequency content and to convert mechanical vibrations to neural spikes. The mechanical movements of the ossicles produce displacement in the cochlear fluids and the basilar membrane. The resulting vibration of the basilar membrane takes the form of waves that travel away from the stapes. For a pure tone, the wave's amplitude reaches a maximum at a certain position along the basilar membrane and then collapses. The position of the maximum amplitude depends on the frequency of the tone; high frequencies have a maximum amplitude toward the stapes and low frequencies toward the apex. Hence the cochlea can be seen as a spectrum analyzer, comprising a set of band-pass filters.
The vibrations of the basilar membrane result in deflection of the sensory stereocilia on top of the hair cells. This deflection opens and closes mechanoelectrical transduction channels, producing a sound-induced change in the current. The change in the voltage across the membrane of the sensory neurons results in the release of neurochemicals and stimulation of neurons in IHCs. The OHCs, on the other hand, the voltage controls a molecular force generation mechanism, also known as the cochlear amplifier. These forces increase the vibration of the basilar membrane.
Auditory nerve
The auditory nerve transmits the spikes generated by the IHCs to the cochlear nucleus. For stimulation with single tones at levels above threshold, the average response rate of the auditory nerve increases roughly linearly with the logarithm of the sound pressure over a limited intensity range. At the onset of a tone, the auditory nerve fibers show initially a very high discharge peak, followed by a lower steady-state discharge rate. The difference between the initial (relatively high) and steady-state rate is referred to as adaptation. This adaptation occurs in three stages: the initial stage has a time constant of a few milliseconds, and is followed by a second stage with a longer time constant in the order of tens of milliseconds. A third, even slower stage lasts tens of seconds.
A very prominent feature for responses to low-frequency tones is the presence of multiple peaks spaced at time intervals corresponding to the period of the stimulus frequency. Thus, the temporal structure of the firing rate reflects the periodicity of the stimulus. This is referred to as phase locking [20, 155, 160, 269]. With increasing frequency, the strength of this phenomenon decreases.
Cochlear nucleus
The auditory nerve enters the cochlear nucleus and divides into two branches: an ascending branch that innervates the anteroventral cochlear nucleus (AVCN) and a descending branch that innervates both the posteroventral (PVCN) and the dorsal nucleus (DCN).
Auditory brainstem
At this and subsequent levels of the auditory system, the information present in the auditory nerve undergoes important transformations. Information from the two ears is integrated to extract binaural cues, and significant monaural features are extracted from the input. The superior olivary complex (SOC) is the first place at which afferents from the two cochlear nuclei converge. In most mammals two major types of binaural neurons are found in the SOC. Both the medial and lateral superior olives (MSO and LSO, respectively) receive binaural input and contain neurons sensitive to interaural time differences (ITDs).
Cells in the MSO receive excitatory input from the cochlear nucleus of both sides and are therefore designated excitatory-excitatory (EE). These cells are sometimes referred to as concidence detectors and their behavior can be compared to a cross-correlator. Their discharge rate in response to binaural stimulation depends on the interaural time difference (ITD) and, at favorable ITDs, i.e. when exhibiting maximum response, typically exceeds the sum of the responses for either ear alone. This favorable ITD is referred to as the cell's best delay. If a given neuron is activated by different frequencies, the different periodic discharge curves appear to reach a maximum amplitude for the same interaural delay of the stimulus. This delay is referred to as the cell's characteristic delay and provides an estimate of the difference in travel time from each ear to the coincidence detector. Conceptually, a set of EE-type neurons with different characteristic delays can be compared to a cross-correlation function [152].
A second subgroup of cells in the lateral superior olive (LSO) and a subgroup of cells in the inferior colliculus (IC) are excited by the signals from one ear and inhibited by the signals from the other ear [7, 8, 33, 158, 175, 189, 204, 224]. The cells in the LSO are typically excited by the ipsilateral ear and inhibited by the contralateral ear and are therefore classified as EI-type (excitation–inhibition) cells. For neurons situated in the IC the excitatory and inhibitory channels are typically reversed and these cells are classified as IE-type cells. The opposite influence of the two ears makes these cells sensitive to interaural level differences (ILDs). With increasing inhibitory level, the neuron's activity decreases up to a certain level where its activity is completely inhibited. The ILD necessary to completely inhibit the cell's response varies across neurons [205, 206, 257]. We refer to the minimum interaural intensity difference needed to completely inhibit the activity as the neuron's characteristic ILD. There are some suggestive data for the LSO [206, 257] and for the IC [133] that the ILD sensitivity of EI-type neurons reflects the differences in threshold between the excitatory and inhibitory inputs that innervate each EI-type cell. In addition to ILD sensitivity, EI-type cells have been reported to exhibit ITD sensitivity as well [157, 158, 205].
It is however uncertain to what extent EI-type neurons contribute to binaural hearing phenomena in humans. It is estimated that ITD sensitive IE units comprise only 12% of low-frequency units in the IC [204]. Furthermore, anatomical studies revealed that the LSO in humans is much less well developed than in various other animals [200].