
Chapter 20
Recent Studies on the Effects of Oil
Abstract
The Deepwater Horizon oil spill in the Gulf of Mexico has stimulated a surge in new research into the effects of oil. Although this new work has largely focused on species, oils, and conditions common to the Gulf of Mexico, many of the lessons can be reasonably extrapolated and applied to other situations and exposure scenarios. A synthesis of the large body of impact studies performed under the Natural Resource Damage Assessment has suggested that many of the effects determined across many levels of biological organization are conserved and common to many different taxa.
Because it is not possible to be comprehensive about the effects of oil in a single book chapter, the discussion here will take advantage of new insights emerging from recent and ongoing work related to the Deepwater Horizon incident. While the chapter will focus on the state of knowledge for only three groups of animals (fish, marine mammals, and sea turtles), the background and science for these taxa provide an excellent overview of research and forensic approaches to oil effects, as well as the status of what we currently understand. The chapter opens with a review of the history of oil effects research to provide context for this complicated topic.
With new research results continuing to be reported in the literature, we can reasonably expect our understanding of oil effects to progress and expand in the near future. For spill responders, the challenge will be to remain informed and translate the new and improved science into oil spill response guidance.
Keywords
Deepwater Horizon; Exxon Valdez; Fish embryos; Marine mammals; Oil effects; Oil toxicity; Sea turtles; Toxicology
20.1. Introduction
It has been previously and frequently noted [1] that the subject of the environmental effects of oil is complex, and a difficult topic to discuss in a way that is both understandable and sufficiently broad enough to convey the range of impacts one may encounter. However, at this point in time, we have learned much and have a more refined and integrated perspective for the ways in which oil affects organisms and populations. Observations and research initiated during and after the Deepwater Horizon oil spill in the Gulf of Mexico have already added to our understanding of the behavior, fate, and effects of oil released into the environment, and so it is an opportune time to revisit the subject of oil effects. Recent findings both build upon previous work provide the basis and direction for the future. The challenge for the oil spill response community is to translate the existing and the new scientific insights into operational guidance for the here and now.
The effects of oil in the environment remains no less challenging a topic than it has been in the past. Recognizing that it would not be possible to write a comprehensive treatise, I chose to frame the present chapter by providing some historical context for how oil toxicity has been considered over the years, and then highlight some of the recent results and observations—some of which, I think, constitute a significant shift in understanding of how oil impacts the exposed organisms. Natural Resource Damage Assessment (NRDA) studies initiated as part of the Deepwater Horizon spill litigation, and ostensibly independent research initiatives made possible by funding from the responsible party, have provided unprecedented opportunities for scientific investigation into short- and long-term effects across many biological levels of organization and a broad range of taxa. There should be findings to digest for years to come.
Those of us who inhabit the world of oil spill response and assessment often rely on case studies as empirical vehicles to illustrate fate and effect, and the same approach is embraced here. Some of the most robust and surprising findings related to oil effects have been those for the early life stages of fish, and on the cetacean marine mammal species documented to have been heavily exposed to oil. The Gulf of Mexico spill was among the first to have affected large numbers of sea turtles, and how these animals responded, both to oil and to our efforts to remediate their exposure, was also enlightening. These examples will be discussed in detail here, with some history of what was known before Deepwater Horizon to provide representative reference points for the evolution of our understanding.
It is an exciting time for people with an interest in oil toxicology; oil spills typically provide the impetus—and importantly, the funding—for research. Interpretation, publication, and synthesis of results from initial and ongoing studies will expand the boundaries of our knowledge and create a solid basis for forecasting and understanding effects during future spill events.
20.2. Some Historical Background
In the first part of the 20th century, petroleum was just beginning its integration into modern industrial life. Beyond its value as a lamp oil that was independent of far-ranging hunting fleets and the dwindling populations of the great whales, petroleum was also found to have a nearly endless set of applications compatible with the emergent machine age. However, its use as a fuel for changing modes of transportation and commerce would make the so-called “rock oil” an increasingly valuable, and ultimately necessary, commodity.
Just as the shift from sail power to coal/steam revolutionized ocean transport in the 19th century, the conversion from coal to oil profoundly changed the history of commerce and engineering in the first decades of the 20th century. In place of coal, heavy fuel oil was used to heat boilers for the production of steam (in 1936, Adam [2] estimated that the British Navy alone used over a million tons of oil per year for vessel steam propulsion), and diesel was increasingly used for internal combustion engine power. Roberts [3] identified 1906 as the turning point for the latter shift, and cited the ease and rapidity of bunkering, increased cargo space, smaller engine staffs, and greatly increased running economy as some of the major advantages in the adoption of oil as a fuel both for steam and for diesel-engine ships. This growing reliance on oil as a vessel fuel was noted by Roberts as a reason for increasing volumes of oil spilled into the sea; he cited an anonymous American oil expert who estimated that 1,500,000 gallons of oil was being discharged daily. Roberts commented [3]:
Of recent years pollution by oil has assumed grave proportions. Oil has fouled beaches and thus destroyed the amenities of many seaside resorts, floating oil-films have been the cause of fires in harbours and innumerable birds have perished miserably through the clogging of their wings and beaks.
The history of our large-scale interaction with oil has been relatively brief, and so it is not surprising that our understanding of its environmental effects is recent as well. Initially, as noted above, the gross effects of combustion and fouling were the obvious consequences of industrial oil use and “routine” discharges into navigable waters. However, a first-person account [4] from what was likely the first large oil tanker spill, the 1907 wreck of the seven-masted Thomas W. Lawson in the Isles of Scilly (off the coast of England, and an estimated two million gallons of paraffinic oil) suggested both human exposure hazards and the toxicity implications of oil spills that would become more familiar with the occurrence of more spills:
The oil floated upon the waves; we could see it washing up here on the shore. At first we could not think what it was—it made the water look black; but soon we learnt from the smell—in fact, we were almost driven away by the smell. They say if it had caught fire it would have cleared the islands, it would have been like a sea of fire, and the smoke would have suffocated all the islanders. As it was, many of the rabbits and birds on Annet were killed by the oil, and lay dead upon the shore.
With growing oil production, transport, and use, therefore, it did not take very long to reveal that there were consequences associated with the convenience of lighting and fuel. One of the earliest scholarly treatises on the adverse effects of oil in the environment was an article [5] by Dr. Emil Seydel in 1912. Seydel described the growing international problem of oil pollution from increasing industrial use, and suggested that an important mechanism of effect was through interference with normal aquatic respiration and oxygen consumption. His observations were surprisingly robust despite representing some of the earliest reported toxicological comparisons in the literature:
It is by direct toxicity, however, that petroleum pollution produces the most severe damage. Naphtha-based substances are toxic to all animal life. This effect can be observed in a simple experiment: If fish are placed in water laced with naphtha, they die sooner than fish in similar water without naphtha. Different kinds of oil have different degrees of toxicity, but (raw petroleum) is the most toxic…Some initially innocuous products of petroleum such as lubricating oil and machine oil acquire greater toxicity under the oxidizing influence of sunlight and oxygen in the presence of water.
Translation from the original German courtesy of R. Childs.
The increasing occurrence of oil releases and growing awareness of impacts led one observer to state in 1925 (cited by Chipman and Galtsoff [6]), that “Oil is, gallon for gallon as thrown out, the most destructive to aquatic life of all the foreign substances now entering our coastal waters.”
Around the same time, Roberts [3] reviewed the existing oil toxicity literature and summarized the findings of both toxic impact and minimal or no impact, with broad variability across different organisms studied. There was recognition of many physical characteristics of oil and its behavior on and in water as variables complicating the assessment of fate and effect. Roberts hypothesized that toxicity might result from the soluble components of oil, and/or emulsification of oil increasing the adverse effects, and set out to test the toxicity of four different oils (two light fuel oils and two heavy boiler-type oils) to brown trout. His results showed that all four oils were toxic to fish. Interestingly, weathering the oils—i.e., exposing them to air and light for 3 months—altered the toxicity results, reducing that of the light oils and not affecting or increasing slightly for the heavy oils.
In 1936, Galtsoff [7] updated what was known about the fate and effects of oil discharged from the sources common at the time (distilleries, refineries, tankers, oil-burning steamships, leakage, and waste from oil fields). Even though the suite of processes we now regard as “weathering” was poorly understood, awareness of some of the subtleties of biological effect did exist:
The natural phenomena responsible for the disappearance of surface films (of oil) are…vaguely understood, and little is known about the eventual fate of oil thrown into the sea. From the biological point of view the disappearance of the oil from the surface does not mean that the pollutant has been eliminated…its effects may still persist when no traces of oil are visible on the surface.
Galtsoff described various ways in which oil could affect marine life, i.e., by interfering with gaseous exchange, by physically coating organisms, by direct toxicity of soluble fractions, by inhibiting normal movements of organisms into streams and bays, and by interfering with food and prey items.
These early studies established some of the fundamental aspects and challenges for oil effects research that are still relevant today: questions of oil miscibility and creating comparable and consistent exposure solutions; acute versus chronic effects and the linkages between them; differential toxicities resulting from different oil types; differential toxicities in different species and life stages; additive or synergistic effects of other factors in increasing apparent toxicity; and translating or extrapolating effects in the field, from actual releases or spills, from laboratory conditions, to name a few.
Others were much less equivocal about the detrimental effects of oil exposure to seabirds. Adam stated [2]:
If there is any quantity of oil on them, they cannot fly, and it is stated that they cannot swim or dive either; they must die of starvation. It is also stated that the oil sometimes penetrates into the down under the feathers next the skin, thereby displacing the air in this down which normally forms a most efficient thermal insulation against cold, and that the temperature of a bird covered with oil may fall many degrees below the normal; they may therefore die of cold.
Lincoln [8] was more succinct:
A bird once oiled is doomed to death…
However, uncertainty remained regarding the amounts of oil that would adversely affect animals besides birds. Adam [2] described the results as “conflicting”:
The general balance of evidence seems to me to point to little, if any, damage with ordinary amounts of petroleum, except occasionally to beds of shellfish between tidemarks. It has been shown that some kinds of fuel oil, in a layer permanently on top of a tank containing fish, will kill the fish in a course of time; also that if water is well shaken up with fuel oil, it may dissolve traces of substances which are toxic to fish. But these experiments refer to a proportion of oil to water which is very much greater than that prevailing on shores subject to occasional pollution by oil.
Many of these challenges and variables were evident in studies reported in the early oil toxicity literature, but evidence continued to mount that oil was not inherently innocuous in the environment. Westfall [9] described toxicity tests using goldfish and oily effluent collected in 1938 from a Socony-Vacuum Oil Company outfall discharging into the Buffalo River in New York. While 10-day exposures to 1:1000 and 1:100 dilutions did not kill test fish, the latter concentration caused fish to become lethargic and to cease feeding. Westfall concluded that the oil exposure caused “disturbances of the central nervous system and of the gastro-intestinal tract.”
In 1961, a classic oil toxicity study by Tagatz [10] exposed juvenile American shad to the water-soluble fractions of three different oil types (gasoline, diesel, and bunker fuel oil), and the median tolerance limit TLm (also known as LC50, the concentration of a substance lethal to 50% of a test population) was calculated for 24, 48, and 96 h. The study found that lighter petroleum products, especially gasoline, were most toxic. Diesel fuel was less toxic than gasoline, and bunker oil least toxic. The 48-h TLm concentrations were 91, 167, and 2417 mg/L, respectively. While Tagatz stated that the study did not determine the mechanisms of the observed toxicity, he noted that the behavior of the exposed fish prior to succumbing (i.e., loss of equilibrium) was consistent with central nervous system disturbance.
The general template of oil effects—that is, lighter oils, with more water-soluble components, being more acutely toxic than heavier oils; heavier oils presenting a physical smothering hazard; and narcosis or desensitization representing a common subacute reaction of exposed organisms—defined much of our understanding of oil toxicity, based on the laboratory studies that had taken place. However, oil effects are not the same as and do not necessarily correlate to oil spill effects.
With ever-larger volumes of petroleum being produced and transported around the globe, spills of oil into the environment became increasingly common. The growing list of such accidental releases represented opportunities to document subtle impacts to less visible and (some would say) less charismatic organisms than seabirds, whose susceptibility had been known for decades. Possibly the earliest oil spill monitored for broader biological impact was that of Tampico in 1957. Wheeler North of the California Institute of Technology recorded the large-scale and mostly indiscriminate initial toxicity of the spilled diesel oil to intertidal organisms (Fig. 20.1). But even more surprising: North [11] continued to revisit the spill site annually through 1984, a period of for 27 years—even now, one of the longest environmental monitoring efforts. His general observations [11] included:
• near-total mortality initially in the vicinity of the wreck;
• no visible oiling at the grounding site after 2–3 months;
• after 2–3 years, effects observed only in longer-lived animals like abalone and sea urchins (i.e., skewed age structure);

Figure 20.1 Fish, crustaceans, and mollusks killed by diesel oil spilled from Tampico, April 25, 1957. Photo: Wheeler North/California Institute of Technology.
• “So after three years, things were pretty normal but they weren't completely normal for quite a long time.”
During the response to the first supertanker spill in 1967, Torrey Canyon, the British government's chief science advisor convened a committee of naturalists and scientists to evaluate the impacts to wildlife communities other than birds, which were being addressed as a separate response. A book published in the same year as the spill described the intertidal impacts as “wholesale slaughter” [12]. However, many of the observed effects were attributed to the detergents applied to the shorelines, not necessarily to the oil. In the case of Torrey Canyon, the proximity of the Plymouth Laboratory of the Marine Biological Association to the spill-affected area facilitated long-term research and monitoring efforts by skilled biologists. Southward and Southward, for example, concluded that recovery of heavily impacted shorelines was still incomplete after 10 years, and observed the importance of biological interactions in determining the structure of recovering intertidal communities [13].
The first modern day large spill in the U.S. was the 1969 Union Oil platform blowout off the coast of Santa Barbara, California. While a large amount of oil (estimated to be 3.3 million gallons) was released and impacts to birds and amenity beaches were highly publicized, impact assessment studies [14,15] did not document large-scale or long-term biological effects.
The range of short- and long-term impacts reported for just these three spill examples illustrates the difficulties inherent in generalizing oil effects. By 1973, a number of large oil spills had occurred worldwide, and more laboratory-based oil effects research had taken place. The National Research Council of the National Academy of Sciences convened a workshop on inputs, fates, and effects of oil on the marine environment, resulting in a summary document [16]. The chapter discussing effects was more of a catalog than a synthesis—perhaps reflecting the challenge of extracting generalities from the empirical field studies and laboratory research that reported a very broad range of results. However, it represented a snapshot of the state of knowledge at the time. It conveyed the realization that the overall biological impact of a spill was not determined simply by toxicity, but by the multitude of variables defining the event: oil type, dosage, oceanographic conditions, meteorology, turbidity, season, biota, and even cleanup. Multiple effects on organisms, communities, and habitats were detailed, but generalizations were scarce beyond the familiar themes of different oils having different toxicities, and different effects with different organisms and different doses.
In 1977, Rice et al. produced a more integrative review [17] of the state of knowledge about oil toxicity that acknowledged the difficulties and contradictions found in the literature, but also provided some explanations or ideas for why such variation was observed, and some recommendations to improve and increase the utility of that research in future. Because Rice et al. were working biologists and chemists, their distilled observations and recommendations were very practical in tone:
Methods
1. Oil toxicity tests require chemical analysis to confirm concentrations of oil in the water.
2. Statistical analysis generating confidence intervals is more informative than graphic estimates of the LC50.
3. If oil toxicities are compared, reference toxicants and the same test species should be used.
4. If species sensitivities are compared, flow-through tests with stable oil concentrations during exposure should be used.
Comparative Toxicity
1. Oil toxicity depends on the concentration of toxic compounds in the oil and physical factors, such as temperature and viscosity of the oil, which affect the transport of petroleum hydrocarbons into the water.
2. Refined oils are more toxic than crude oils on a volume-added basis because refined oils often have higher concentrations of toxic compounds.
3. The toxicity of oils is apparently due to water-soluble compounds rather than dispersed droplets.
4. The toxicity of aromatic hydrocarbons increases with the number of rings and with the degree of alkyl substitution. However mono- and di-nuclear aromatics account for most of the toxicity in water-soluble fractions.
Comparative Sensitivity
1. No conclusions can be made concerning egg sensitivities because not enough quantitative data exist.
2. Crustacean larvae appear more sensitive than most adults. Molting animals are more sensitive than non-molting animals.
3. Sensitivity data for larvae or juveniles from other groups are generally lacking, except for polychaete juveniles which are more tolerant than adults.
4. Adult sensitivities are fairly similar when data from static tests are compared. Intertidal animals appear more tolerant in short-term, static exposures, probably because of their ability to temporarily insulate themselves from the environment.
5. Cold-water species probably have sensitivities equivalent to the sensitivities of warm-water species, but cold-water species may be more vulnerable to oil toxicity because toxic aromatic hydrocarbons persist longer at colder temperatures.
Recommendations for Future Research
1. Standardized methodology (flow-through tests, chemical analyses, and statistical analyses) should be used in future toxicity tests.
2. There is little need to test the toxicity differences between different oils. Future research should:
a. determine if aromatics account for most, if not all, of the toxicity of water-soluble fractions (WSF), and
b. determine the relative importance of individual mono- and di-nuclear aromatics to the acute toxicity under various conditions.
3. Additional sensitivity data are needed for eggs, larvae, and juveniles of several groups of animals such as mollusks, echinoderms, etc., since virtually no information on these animals is available.
4. Several species need to be retested with flow-through exposures, so that
a. better estimates of sensitivity can be measured, and
b. sensitivities can be compared among species.
5. Selected species should be tested for longer periods of time, so that the relationship between oil concentrations that are toxic for short and long exposures can be determined.
Oil effects research continued to progress beyond basic dose-effect studies to eliciting a greater understanding of mechanisms that might provide more practical applied capability. That is, if we know the chemistry of a spilled product, can we predict an expected impact to a given resource of concern? The growing knowledge base and the generally accepted ideas about oil toxicity formed the basis for increasingly sophisticated approaches to effects modeling [18,19] and prediction of impact based on chemical structures and relationships [20]. The concepts of aqueous solubility of the constituents of complex mixtures, and resultant exposure impacts on the function of cell membranes and other tissues leading to narcosis, formed the predominant basis for considering oil toxicity into the 21st century. Di Toro et al. [20] presented a complex model of crude oil toxicity that incorporated the physical chemistry and behavioral characteristics of complex mixtures of hydrocarbons, the tendency of aromatic hydrocarbons and other organic compounds to associate with higher lipid concentrations in biological resources, and the known information about acute lethality of chemical constituents.
A shift in the way the effects of oil and oil spills were assessed and considered occurred around 1989 Exxon Valdez oil spill in Prince William Sound, Alaska. This was not a coincidence, but rather, a confluence of several factors including the environmental persistence of weathered oil residues from the spill incident, the apparent long-term impacts to resources of ecological and commercial significance, the regional presence of a skilled science laboratory, and personnel allocating time and effort to the subject, and lastly, the new inevitability of litigation-fueled scientific disagreements. As we have related, much of the prior toxicological attention for oil effects was targeted on short-term, acute (i.e., lethal) effects of the soluble fractions of an oil mix, i.e., LC50 studies. This applied not just to oil studies, but was the norm across the field of ecotoxicology as a whole. In a review of the state of this field, Brander et al. [21] acknowledged the “simplistic” focus, but recognized it as the bedrock of ecotoxicology in the latter half of the 20th century. It is the progression upon and beyond the long-held fundamentals of oil effects that characterize the science today, presenting both promises and challenges for the response community.
20.3. Oil and Fish: A Paradigm Shift
In 1962, Thomas Kuhn published a book, The Structure of Scientific Revolutions. The premise of Kuhn's book [22] was that the advancement of scientific knowledge was not, contrary to popular belief, by the steady accumulation of information and theory; rather, he proposed that science progresses when revolutionary ideas jolt the established thinking forward into a new, uncharted territory. Key to Kuhn's model was that unexpected outliers and anomalies lead to new paradigms of thought that ultimately change what is accepted as scientific fact.
The notion that oil spilled in the environment conferred its toxic effects quickly, when degree of contamination was extremely high, was consistent with many of the observed impacts during actual oil spills: mass mortalities of birds and other exposed organisms became hallmarks of oil spill impact, with dead and impaired wildlife the most compelling and emotionally evocative indicators of the consequences of oil exposure. Laboratory studies confirmed the obvious: the petroleum mixtures contained toxic components, and oil spills provided apparent empirical evidence that environmental concentrations attained sufficiently high levels to cause direct mortalities. However, integrating or linking the laboratory with the field proved to be challenging; some in the response community minimized potential environmental effects concerns as recently as the 1990s [23]:
The measured field exposures to…hydrocarbons from untreated and chemically dispersed crude oils are much lower than those observed to kill a wide range of organisms in laboratory bioassays…Thus, oil spills and chemical dispersion of crude oil slicks are unlikely to have an adverse impact on larval, juvenile or adult organisms in the water column.
The Exxon Valdez spill represented a major inflection point for how spill effects and spill response itself were defined and perceived in the US. The research initiated mid- or post-Exxon Valdez marked the beginning of a change in how oil spill effects were measured and perceived. Peterson et al. [24], in their synthesis of results from Exxon Valdez studies sponsored by the state and federal trustees, characterized an older paradigm of oil effects defined by laboratory toxicity tests with direct mortality endpoints and the assumptions that an oil spill was a short-term event characterized by rapid decrease in exposure with toxicity primarily caused by water-soluble fractions of oil. They contrasted this now familiar template with an evolving perspective of a more complex and holistic view of impact based on ecotoxicology, potential long-term persistence of oil, chronic effects endpoints, and specifically: larger hydrocarbons as significant drivers of toxicity. This new perspective represented a new paradigm, as Thomas Kuhn would define it, and has reconfigured how oil toxicity to fish is understood.
In 1977, researchers from the NOAA/National Marine Fisheries Service laboratory in Auke Bay, Alaska, summarized what was known about oil toxicity. Their insights [17] were discussed above. Two decades later, Rice et al. [25] described their nonlinear investigatory path from 1977 to the post-Exxon Valdez years, showing that it was an iterative process that began with what was believed to be known about oil fate and behavior, and validated (or not) against the reality of conditions in Prince William Sound. For example, Rice et al. evaluated measured concentrations of polynuclear aromatic hydrocarbons (PAHs) under the oil slicks and initially concluded [25] that the environmental levels were too low to be toxic:
… the low concentrations of oil in the water under the slicks were presumed to be too low to be acutely toxic, because the concentrations were substantially below the acutely toxic levels reported in the literature of the 1970s and 1980s. For example, laboratory toxicity tests with WSF (water-soluble fractions of oil dominated by 1- to 2-ringed aromatic hydrocarbons) had 96-h LC50 of WSF for pink salmon ranging from 1500 to 8000 ppb (parts per billion), depending on life stage, salinity, and type of exposure…After the spill, environmental samples had PAH concentrations well below those concentrations found to be acutely toxic during 28-day exposures of most marine organisms to water-soluble fractions…Evidence of acute toxicity from herring larvae field studies (abnormalities) occurred…and it was realized that the composition of the PAH after the spill was substantially different than the WSFs reported in the literature.
The referenced herring larvae research was that of Brown et al. [26], who focused on the 1989 year class of Pacific herring (Clupea pallasi) that hatched and developed immediately after the spill. Herring were a particular concern because they spawn in the nearshore region, and the timing unfortunately coincided with the oil spill. Brown et al. reported that while in situ mortality of embryos incubated at oiled sites was not found, a number of sublethal effects were determined. These included premature hatch, low weights, reduced growth, and increased morphologic and genetic abnormalities.
The water column chemistry results from oiled waters of Prince William Sound showed PAH concentrations to be well below the “classical” LC50 concentrations in the literature. But these benchmark acute toxicity concentrations were based on the water-soluble fractions of oil mixtures, primarily the smaller one- and two-ringed PAHs. Three- and four-ringed PAHs, while much more inherently toxic (Rice et al. cited experimental evidence that each additional ring in a PAH structure increased toxicity by an order of magnitude), were also much less water-soluble and thus not regarded as the oil exposure risks. Actual environmental sampling, however, showed low levels of monoaromatics, but “significant” contributions of three- and four-ring PAHs. One emerging bottom line: the exposure assumptions for many, if not most, of the available acute toxicity studies were not particularly relevant to the conditions occurring after the Exxon Valdez spill, and empirical field results suggested that there were other factors in play.
The realization that—contrary to most of the assumptions from the preceding six decades—a chemically different set of petroleum components might be significant toxicological drivers rather than what had been assumed, carried with it a number of potent implications. For one, the basic understanding of how oil weathered, that the lighter hydrocarbons (e.g., one- and two-ringed PAHs) were relatively short-lived in the environment, had reinforced the notion that oil spill impacts were short-term and quickly diminishing with time. However, larger hydrocarbons were known to be more environmentally persistent [27]; if these were the critical drivers of toxic effect, then much of what was believed about oil toxicity would require some recalibration.
Pacific herring were not the only species of commercial importance that was judged to have been especially at risk from the Exxon Valdez spill. Pink salmon (Oncorhynchus gorbuscha) were the dominant commercial fisheries harvest in Prince William Sound, and in contrast to other salmon species, spawn in the shallow intertidal mouths of streams and rivers—where they were at risk from potential exposure to nearshore and stranded oil on beaches. As research subjects, pink salmon had the added advantage of having been studied for many years as an important commercial fishery species in Alaska, and particularly Prince William Sound.
Carls et al. [28] collected juvenile pink salmon and chum salmon (Onchorhynchus keta) from oiled and unoiled areas of Prince William Sound in the first two years of the spill, 1989 and 1990. They found significantly higher concentrations of PAHs (i.e., 44 parent and alkylated homologs of two- to five-ring PAHs and dibenzothiophenes) in salmon tissues in 1989 (181 ± 28 ng/g vs. 54 ± 8 ng/g), but not in 1990. The PAH composition found in salmon tissues was similar to that of the original source oil. The cytochrome P4501A enzyme system, the activation of which was recognized as an indicator of exposure to organic hydrocarbons including PAHs, was also significantly induced in oiled areas in 1989.
As was the case with Pacific herring, direct mortality of adult salmon was not reported in the field, indicating that acute toxicity levels had not been attained. However, endpoints such as reduced larval growth and increased incidence of developmental abnormalities indicated the occurrence of sublethal effects of exposure to oil, and Bue et al. [29] reported higher pink salmon embryo mortality at oiled beaches than reference beaches. Building on field observations of higher than control salmon embryo toxicity at oiled sites in Prince William Sound, Heintz et al. [30] continued the NOAA pink salmon studies by exposing embryos to very low concentrations of weathered oil in controlled, but simulated field conditions. They determined that mortality rates for embryos exposed to PAH concentrations derived from very weathered (for 1 year) oil, as low as 1.0 part per billion, were significantly higher than those from less weathered oil.
The initial conclusions from the Exxon Valdez herring and salmon studies were not without controversy and were not universally embraced. The Exxon Valdez spill inspired and defined new norms and practices for oil spill preparedness, response, and research; one outcome was the emergence of adversarial science whose outcomes would be determined by courts of law, and not necessarily by peer review. With billions of dollars in civil and criminal penalties potentially at risk, this was probably inevitable.
Brannon et al. [31] conducted a separate set of pink salmon studies that arrived at distinctly different conclusions than did federal and state studies. They established experimental locations in the field, defined as oiled or reference, and then allowed fertilized salmon eggs to incubate in situ. They concluded:
The early life-history studies we conducted…demonstrated that the problems that might have been produced by the oil spill, in fact, did not occur…Whether these data are interpreted to mean that pink salmon were not affected by oil, or alternatively, pink salmon must not have been exposed to biologically meaningful concentrations of oil, the result is the same. There was no detectable effect of the Exxon Valdez oil spill on the (Prince William Sound) wildstock pink salmon population.
In their review [25], Rice et al. acknowledged both the controversy and the scientific disagreement, and for each endpoint type attempted to reconcile why state and federal trustee scientists arrived at one set of conclusions, while Exxon researchers arrived at a very different set. They attributed some of the differences to sometimes subtle differences in sampling strategies or methodologies, but in the end, they noted reconciliation was not always possible:
In some instances, these studies agree, but in several cases, especially the areas involving effects on various life stages, similar studies, but with different experimental designs, have led to differing findings and conclusions. Studies have examined both short- and long-term effects, and disagree on the extent of the damage, and even on the existence of damage to pink salmon. Several studies examine the same issue but were seldom conducted identically. Consequently, most of the research effort is more (complementary) and open to differences in interpretation.
Carls et al. [32] synthesized information about the toxicology of Exxon Valdez oil to Pacific herring, and similarly acknowledged the scientific disagreements that had come to characterize aspects of the environmental studies related to the spill. Their perspective was that reconciling the differing conclusions was hampered by differences in methods for oil detection, estimating the size of the spill itself, mechanisms of oil toxicity, and interpretations of population data. Nevertheless, in contrast to the situation for pink salmon, there was actually some degree of consensus between both groups of scientists, and they agreed that by 1990, oil impacts to both embryos and larvae could not be detected. However, the situation with Pacific herring became more complicated in 1992–93, when Prince William Sound stocks collapsed (according to Elston and Meyer [33], a 63% decline from expected returns) and closed the multimillion-dollar commercial roe fishery.
Carls et al. [32] had noted that incidences of premature hatch and skeletal, craniofacial, and finfold abnormalities was higher at oiled sites than unoiled sites. Carls et al. [34] linked sub-parts per billion concentrations of PAHs to similar malformations, genetic damage, and mortality in Pacific herring embryos. Marty et al. [35] showed that water concentrations of total PAHs as low as 4.4 parts per billion resulted in significant biological effects (e.g., induction of the cytochrome P4501A mixed function oxidase enzyme, development of ascites (fluid in peritoneal cavities), and increased mortality) in pink salmon larvae. Low-level exposures of crude oil to eggs and larval fish resulted in consistent and distinctive deformities including (but not limited to) craniofacial abnormalities, spinal curvature, malformed and misshapened fins, impaired eye development, and yolk-sac issues (Fig. 20.2), reflecting some convergence between field observations during the spill with laboratory results from several studies.
However, an unanswered question and unresolved issue is whether and/or the degree to which the Exxon Valdez oil spill was responsible for the collapse in the herring populations in Prince William Sound. Carls et al. [32] did not find long-term effects to herring, but would not rule out indirect linkages. When the Deepwater Horizon spill occurred, media and popular opinion referenced the Exxon Valdez and herring, and attributed oil exposure as the primary cause of the fishery collapse. However, the science is more equivocal and uncertain. Infection of the herring population by viral septicemia virus was, for many years, thought to be the leading potential cause; but Elston and Meyers [33] reviewed several key studies and concluded that it was not. Furthermore, they investigated whether exposure to weathered crude oil correlated to increased susceptibility to the viral infection, and after extensive review, failed to confirm a link. Elston and Meyers presented an alternative and multifactorial hypothesis: that while the virus may have played a secondary role in the herring population crash, they believed that body condition of herring had in fact been declining for eight years beginning in the mid-1980s. While the population biomass peaked in 1992, food availability was poor and fish lost weight over the winter when the collapse occurred. Moreover, the crash encompassed all year classes and was not age-specific—which would have been expected with a viral infection. They suggested that the weakened population of herring died off during the winter of 1992 from predation and other “natural” causes.

Figure 20.2 Normal herring embryo (top); oil-exposed embryo (bottom) showing typical spine curvature, cranial malformation, and edema. Photo: Mark Carls/NOAA.
Investigating the biochemical mechanisms behind the growing weight of evidence indicating a toxic syndrome of oil exposure causing developmental abnormalities in larval fish, Barron et al. [36] used the toxicity and PAH chemistry results from Carls and his colleagues in Auke Bay to evaluate the performance of existing PAH toxicity models (narcosis, aryl hydrocarbon receptor (AHR) agonism, alkyl phenanthrene toxicity, and combined toxicity that assumed additive toxicity from the three mechanisms). Their analysis showed the highest degree of correlation between the observed embryonic toxicity results and the alkyl phenanthrene model that was based on reports [37,38] of significant early life stage effects to fish embryos exposed to alkylated phenanthrenes.
Incardona et al. [39] at the NOAA/Northwest Fisheries Science Center studied the physiological mechanisms underlying the observed effects of oil exposure to embryos of the freshwater zebrafish (Danio rerio), a robust aquatic test organism. They assessed the differential toxicities of seven PAH parent compounds (naphthalene, fluorene, dibenzothiophene, phenanthrene, chrysene, anthracene, and pyrene) to zebrafish embryos. The first five PAH compounds were chosen to provide a degree of comparability to those found in weathered Alaska North Slope crude, the oil spilled from the Exxon Valdez, while the last two were much less prevalent but with similar ring structures and were included to provide insights into potential structure–activity relationships. No gross anatomical deformities resulted from exposure to naphthalene, anthracene, or chrysene. In contrast, larvae exposed to fluorene, dibenzothiophene, and phenanthrene developed dorsal curvature of the trunk and tail, and growth reduction of the head. Moreover, treatment with dibenzothiophene and phenanthrene, both PAHs with three-ring structures, resulted in severe pericardial and yolk-sac edema.
The AHR is an important regulator in physiological and pathophysiological processes, including xenobiotic metabolism, endocrine function, immunity, and cancer. Agonist activation of the AHR is considered deleterious based on toxicological evidence obtained with environmental pollutants (particularly dioxins), which mediate toxic effects through AHR and activate the CYP1A1 enzyme system, the familiar biomarker for hydrocarbon exposure. PAHs are known to bind with the aryl hydrocarbon receptor to initiate the production of enzymes to convert the PAHs to more water-soluble metabolite to facilitate excretion. However, AHR activation may also result in a toxic response if the binding ligand (i.e., the contaminant) is not a good fit. In this case, as explained by Incardona et al. [40], the toxic substance builds up in tissues and continually activates the AHR. In addition, the potential for creation of reactive and detrimental intermediate metabolites also exists during the detoxification pathway.
Although Barron et al. [36] had inferred a general model of PAH toxicity that might explain the underlying physiological mechanism behind the larval fish impacts, Incardona et al. [40] investigated AHR agonism and whether zebrafish AHR activation by dioxin was the same as that by three- and four-ring PAHs. Unfortunately, for those who might have hoped for some simplicity deep in the midst of a complex topic, Incardona and colleagues found that not only did AHR activation differ for dioxin and PAHs, but also different PAH types acted by different toxic pathways during larval development. Moreover, even though chrysene was found to be generally nontoxic to zebrafish embryos, it nevertheless activated AHR in different tissues and was a potent CYP1A1 inducer.
However, the similarity of results obtained when larval fish were exposed to weathered crude oil and to the three-ring PAHs, strengthened the notion that the tricyclic components of crude oil comprised the most toxic fraction. Incardona et al. concluded that the broader implications of this work included:
• importance of understanding the chemistry of oil spilled in the environment;
• affirmation of cardiovascular dysfunction as a cause of the observed deformities in exposed embryos;
• their research indicated that the widely used marker of PAH exposure, induction of CYP1A1, may reflect its role as a protective mechanism rather than a cause of oil-related toxicity; and
• realization that relatively low molecular weight but relatively ubiquitous three-ring PAHs such as phenanthrene conferred greater toxicity than had been previously modeled.
While previous work had suggested that exposure to complex petroleum mixtures, such as weathered Alaskan crude oil caused these kinds of developmental issues and deformities, researchers had narrowed the classes of compounds (i.e., three-ringed PAHs) that seemed to be responsible, and began to identify the physiological mechanisms responsible. In particular, they suspected cardiac dysfunction resulting from exposure to the three-ring PAHs as the key interaction driving impaired development of fish embryos. They were able to strengthen that argument by comparing the consequences of PAH exposure to those from treatment with a known genetic disruptor (“gene silencing”) of cardiac function and found the results (dorsal curvature of the spine, impaired eye and jaw development) to be essentially the same.
The sum of this work with larval fish, first in Alaska in the wake of the Exxon Valdez, and then in subsequent studies that built on the spill studies and then focused on physiological and subcellular mechanisms, described a fundamentally different paradigm of oil toxicity than had been the norm for the preceding decades. This was recognized by the researchers themselves, some of whom having been instrumental in establishing the first paradigm. Rice et al. [25] summarized the old way of thinking about oil toxicity in the following ways:
• Focus was on one- to two-ringed PAHs, acute toxicities, and short-term (e.g., 96-h) exposures.
• Comparison of toxicities was among oils, sensitivities between species, or life stages.
• Although increasing toxicity of the three- to four-ringed PAHs was known, the presumption was that there was little bioavailability of these compounds following a spill.
A key part of the paradigm shift was the realization that persistence of residual oil in the environment resulted in a qualitatively different chemical exposure scenario than what had been almost taken for granted in laboratory testing for decades. The long-term persistence diminished the toxicological influence of the smaller aromatics, but emphasized or exaggerated that for the larger, more inherently toxic, but less bioavailable PAHs. In addition, the fish embryo studies consistently showed that the three- and four-ringed PAHs conferred their toxicity at much lower concentrations than had ever been considered for the standard LC50 studies that had shaped our thinking about the effects of oil for such a long period of time.
20.4. Fish Studies Post–Deepwater Horizon
The current state of knowledge about the effects of oil and specific classes of PAHs to the early life stages of fish can be considered as relatively robust—at least, as Scholz and Incardona [41] qualified, at the scale of the individual animal. The crude oil toxicity syndrome (as they term it) has been well established for several species and different oils, and is consistently characterized by defects in heart formation and function, resulting in craniofacial and finfold abnormalities. Studies with individual components of complex oil mixtures continue to attribute the toxicity primarily to three-ring PAHs such as phenanthrenes, dibenzothiophenes, and fluorenes. The weathering of crude oil naturally enhances or enriches the relative proportion of the three-ring aromatics in the mixture by selectively reducing the concentrations of lighter hydrocarbons. While the triaromatics tend not to be very water soluble, they have been shown to confer their toxicity at very low concentrations.
With the toxicity syndrome fairly well described and validated, the challenge now is to address the “So what?” question: that is, what are the implications of these early developmental effects to individual fish, to their populations, and ultimately, to communities and ecosystems? Would the pathologies observed in the laboratory be expected to occur at chemical concentrations typically encountered in field exposures? Do abnormal hearts and morphological deformities translate into impaired performance of individuals, and then to the other larger levels of biological organization? Walker et al. [42], in a basic text on ecotoxicology, featured a simple graphic (Fig. 20.3, below) that shows the different levels of biological organization where we might consider effects of a contaminant. It also illustrates the quandary researchers—and environmental responders—face in interpreting biological effects: that the organizational levels to which we typically assign the greatest importance (i.e., population, community, ecosystem) require the longest time to respond to exposure, and are most difficult to link back to a particular chemical. Exposure to chemicals found in oil present another challenge, in that the nature of exposure is both dynamic (potentially changing dramatically with time) and expected to be transient (diminishing as weathering and cleanup take place).
Particularly with respect to the implications of exposure across different levels of biological organization, ecotoxicologists understand this. For example, Scholz and Incardona [41] commented:
The current state of the science is now focused on linkages across biological scales, from channel blockade and the disruption of excitation–contraction coupling in individual heart muscle cells to heart development, circulatory and respiratory performance, delayed mortality of individual fish, and associated consequences for the abundance and dynamics of wild populations. For forage fish such as Pacific herring, the biological scaling will eventually extend to communities and ecosystems.

The Deepwater Horizon oil spill provided the need and motivation to further validate the oil and PAH toxicity model that had emerged from the Exxon Valdez and had been refined through studies with tropical freshwater fish—but with species of concern from a completely different marine environment: the Gulf of Mexico. Incardona et al. [43] bridged between Exxon Valdez and Deepwater Horizon, and set the stage for new studies to validate and build upon the cardiac toxicity paradigm of oil effect in fish. They again used the freshwater zebrafish (D. rerio) to compare the effects of the two oils (Alaska North Slope crude, and fresh and weathered Mississippi Canyon 252—MC252—oil that was released during the Deepwater Horizon) from the two iconic spills, and determined that same cardiotoxic pathologies were caused by exposure to the two oils.
Precisely how those cardiotoxic pathologies were physiologically conferred is not nearly as well elicited, but recent work in the wake of the Gulf spill provided new insights here. Brette et al. [44] used cardiac muscle cells, or cardiomyocytes, from juveniles of two pelagic species, the bluefin tuna (Thunnus orientalis) and yellowfin tuna (Thunnus albacares) to examine the physiological mechanisms causing the cardiotoxic effects. They focused on the biochemistry of muscle contraction, the excitation–contraction coupling, which is the process by which an electrical stimulus is converted into a mechanical response—in this case, contraction of heart muscle. They found that MC252 oil, and particularly, the weathered MC252 oil, interfered with that excitation–contraction coupling in the tuna cardiomyocytes through disruption of potassium and calcium ion functions at the cellular level of developing hearts. The fact that the effect was enhanced with weathered oil supported the earlier findings that the more persistent three-ring PAHs, enriched by weathering, were the components of oil driving this effect.
Because many pelagic fish species spawn in the Gulf of Mexico during the seasonal time frame when oil was released during the Deepwater Horizon blowout, Incardona et al. [45] investigated the effects of field-collected oil from the spill on embryos of three pelagic warm-water fish, the southern bluefin tuna (Thunnus maccoyii), yellowfin tuna (T. albacares), and yellowtail amberjack (Seriola lalandi). The spill-affected waters of the Gulf of Mexico included known spawning grounds for a number of pelagic fish species whose eggs float near the water surface. For example, highly prized Atlantic bluefin tuna (Thunnus thynnus) numbers have declined considerably and are under consideration for listing under the US Endangered Species Act. The western population spawns in the Gulf, and the timing (April–June) in 2010 coincided with the peak of the spill.
In this study, Incardona et al. found that environmentally realistic exposures (1–15 ppb total PAH) caused specific dose-dependent defects in cardiac function in all three of the test species, with circulatory disruption resulting in pericardial edema (fluid in the sac around the heart) and other previously described typical malformations. Each species displayed an irregular cardiac arrhythmia following oil exposure. Concentrations as low as 2–3 ppb summed PAHs caused edema in more than 50% of tuna embryos. Atrial arrhythmia was the most sensitive indicator of cardiotoxicity, with thresholds near 1 ppb for yellowfin tuna and amberjack. Although the general suite of effects were remarkably consistent not only for these Gulf pelagic species (Fig. 20.4), but also compared to Exxon Valdez-derived results for salmon and herring, small variations across species were attributed by Incardona et al. to such factors as egg size and differences in metabolic capacity during development.
Mahi mahi (Coryphaena hippurus), also known as dolphinfish or dorado, is a popular excitation–contraction recreational and commercial species in the Gulf of Mexico. It is another pelagic fish whose spawning would have overlapped temporally and geographically with the spill. As a research subject, mahi mahi has the advantage of being a rapidly growing species that also can be spawned readily in research facilities. Edmunds et al. [46] exposed mahi embryos to Deepwater Horizon field–collected MC252 oil samples. They noted that previously, the cardiotoxicity syndrome had been assessed visually by means of microscopy; this had been effective, but not completely practical for field-collected or large numbers of embryos, nor for impacted individuals that did not display overt signs of exposure (e.g., edema or craniofacial abnormalities) at the time of assessment. Edmunds et al. sought to identify and compare potential molecular markers of cardiotoxicity with the conventional visual markers. They found that edema was the main morphological response to oil exposure, but interestingly, did not manifest in the development of craniofacial, finfold, and body curvature deformities that were characteristic in many other fish species studied. Ten gene responses related to cardiac development and function1 were evaluated as candidate molecular indicators of toxicity as well as five others2 that were linked to more general stressors. The prospective cardiotoxicity genes were downregulated in nearly all oil exposures. The largest responses were obtained with genes related to myofiber structure and contractility. Several genes diagnostic of cardiovascular stress in humans were also concentration-responsive to oil exposure in mahi mahi. Relative to the visible markers of cardiac injury, the molecular indicators became relatively more sensitive as larval development progressed. The visible measures were more diagnostic in the earlier developmental stages of mahi mahi.

Figure 20.4 Comparison of oil effects in three Gulf of Mexico pelagic fish species embryos: (A, Aʹ) bluefin tuna, (B, Bʹ) yellowfin tuna, and (C, Cʹ) amberjack. White arrows point to heart; black arrows show margins of yolk sac; asterisks indicate absence or deformity of eyes. Photos: John Incardona/NOAA; previously published in Incardona JP, Gardner LD, Linbo TL, Brown TL, Esbaugh AJ, Mager EM, Stieglitz JD, French BL, Labenia JS, Laetz CA, Tagal M, Sloan CA, Elizur A, Bennett DD, Grosell M, Block BA, Scholz NL. Deepwater Horizon crude oil impacts the developing hearts of large predatory pelagic fish. Proceedings of the National Academy of Sciences 2014;111(15):E1510–8.
While Edmunds et al. sought molecular markers of oil effect to mahi mahi, Mager et al. [47] looked in the opposite direction with respect to levels of biological organization to link embryonic cardiotoxicity to individual performance of exposed fish. They exposed mahi mahi embryos to MC252 oil (1.2 ± 0.6 ppb summed PAHs for 48 h) that were then raised for 25 days to juveniles, and also directly exposed juveniles (30 ± 7 ppb for 24 h). Although no typical visible signs of cardiotoxicity (i.e., edema and decreased heart rate) were documented, swimming performance in the exposed larvae and juveniles was significantly impaired (37% and 22% reduction, respectively). The authors implied that survival to maturity for oil-exposed fish would be impacted by swimming performance limitations imposed on key behaviors like foraging, predator avoidance, and migration.
Another variation on this increasingly established theme of oil embryotoxicity to fish was described by Jung et al. [48], who exposed two larval fish species (spotted sea bass, Lateolabrax maculates, and olive flounder, Paralichthys olivaceus) to Iranian heavy crude oil in order to determine interspecific differences in effect (defined as the genesis of morphological defects, uptake of PAHs, and expression of CYP1A). The PAH concentrations causing effects were similar to other fish embryo studies, ≤10 μg/L total PAH. Consistent with the other previously described fish embryo studies, oil exposure resulted in the typical suite of morphological defects, including pericardial edema, reduced finfolds, spine curvature, and delays in development. Generally, the flounder embryos were more affected than the sea bass, with greater degree of defects and developmental delay, greater uptake of PAHs, but curiously, lower CYP1A induction. The authors suggested that the higher CYP1A levels in the sea bass may have reflected a greater degree of biotransformation of PAHs, resulting in the lower body burdens of PAHs in that species. The differences in species effects, while suggesting species differences in toxicokinetics and toxicodynamics, still supported the notion that the fish embryo toxicity paradigm was conserved across a broad range of both fish and oils.
In the years following the Deepwater Horizon spill, the Gulf of Mexico Research Initiative—the $500 million, 10-year competitive grants program established by British Petroleum to fund basic oil effects research—will support further research to understand both the mechanisms of oil toxicity and their implications to individual organisms, populations, and ecosystems. This should provide greater insight into the true environmental costs of oil pollution, but perhaps less guidance on what we can do about it either as responders or as a society with a continued reliance on petroleum. The challenge for the scientists conducting spill-related research under this and other programs will be to provide context that will make their results both relevant and applicable for response and for environmental decision-making in society at large.
20.5. Marine Mammals and Oil: Early Studies
As the preceding narrative reflects, much has been learned about the effects of oil on the early life stages of fish. While illuminating, this is obviously only a small part of the story of oil effects. Very early into the response for the spill in the Gulf of Mexico, concerns were raised about potential exposure to populations of marine mammals that inhabit the area. NOAA responders commissioned aerial overflight surveys in areas with surface oiling to ascertain if marine mammals were present, and it very quickly became evident that they were (Figs. 20.5 and 20.6).
Thirty-one species of marine mammals are known from the Gulf of Mexico [49]. However, little empirical information existed concerning the effects of these kinds of exposures to these animals, including cetacean species observed on Deepwater Horizon overflights to be occurring in oiled portions of the Gulf. Even stock assessments and population surveys can be challenging for marine mammals, as they are obviously mobile and their vertical ranging through the water column makes accurate identifications and counts difficult from the surface or the air. Moreover, the protected status of most marine mammals renders toxicological studies impractical if not impossible.


Engelhardt [50] reviewed the available information with respect to oil and marine mammals and concluded that “At the present level of knowledge, prediction of the effects of petroleum on a given species will require much correlation and inference from what is known about the natural history and physiology of the species, and about the characteristic toxicities of oil in mammalian systems.” A more comprehensive compilation of information on oil and marine mammals was the 1990 book, Sea Mammals and Oil [51], edited by Joseph Geraci and David St. Aubin. In this book, Geraci authored a chapter on physiologic and toxic effects of oil on cetaceans [52] and summarized the known impacts from oil spills to that date. Although a number of incidents intersected with or involved marine mammals (cetaceans), almost no direct impacts could be linked to oil exposure, leading Geraci to conclude that “…there is no gripping evidence that oil contamination has been responsible for the death of a cetacean.”
Geraci also summarized the results of experimental studies he conducted with colleagues [53,54] to determine if captive bottlenose dolphins (Tursiops truncatus) were capable of detecting oil on the surface of the water in their tanks. They concluded that not only could the dolphins detect oil day or night, they learned to avoid it and were subsequently averse to contacting it. Geraci suggested that there was a tactile aspect to oil that was a factor in dolphins' ability to detect and avoid. St. Aubin [55] contrasted the inherent ability of dolphins to detect surface oil in thicknesses from 1 mm to 1 cm, to empirical observations of dolphins and other cetaceans not avoiding oil. He speculated, without further elaboration, that in the wild, dolphins may be reluctant to avoid a spill “…because of some overriding attraction to the area.”
The effects of surface contact with oil, when it did occur, were inferred from human effects and then tested on four species of odontocetes (toothed cetaceans) [54]. The reactions elicited from the marine mammals were much less severe than those noted for humans. Remarkably, even a 16 h exposure to gasoline did not affect the lipid concentration in white-sided dolphin (Lagenorhynchus acutus) skin. Noting that an exposure in an actual spill scenario was likely to be much less extreme than the experimental conditions, Geraci inferred that real-life contact with oil “…would be less harmful than we and others had proposed.”
Geraci discussed other potential routes of exposure and impact to cetaceans, such as inhalation, fouling of baleen plates in mysticete whales, and ingestion. Although he acknowledged vapor inhalation as a real threat to the health of marine mammals, no cast studies or clinical examples were cited. There were experiments in which baleen was soaked in different oils or fouled with oil and then tested for functional impairment, but the results did not, in his opinion, warrant concern. Similarly, while speculative scenarios would suggest that cetaceans could ingest potentially harmful quantities of oil during a large release, little experimental or empirical information exist. One study [56] was cited in which small quantities of oil were fed to a dolphin over a 14-week period with no discernible effect. Geraci felt that ingestion of potentially harmful quantities of oil was unlikely to occur and alluded to the lack of evidence from either observational or stranding studies.
Overall, Geraci did not think it likely that populations of cetaceans would be adversely affected by an oil spill, while acknowledging that individuals could be. He hedged this conclusion by noting some habitats (e.g., the Arctic) were inherently more sensitive; while others (e.g., coastal oil production areas) were inherently more stressed. Nevertheless, very few concerns were identified in this analysis.
Some early assessments of impacts to marine mammals during actual spills seemed to support the notion that cetaceans would be able to avoid oil—even a release that ranks as the largest marine spill in history, the 1991 Gulf War spill. While acknowledging the scarcity of basic and baseline data on marine mammal populations in the affected area, Robineau and Fiquet [57] assessed the population status of one group of humpback dolphins (Sousa chinensis) in two bays and found no evidence of abnormal mortality. Further, they inferred that residents of the bays were able to avoid contact with the oil. However, the assessment and conclusions were based on very limited survey data with few marine mammal sightings.
The 1989 Exxon Valdez oil spill provided the backdrop and the setting for more empirical information on risks and effects to marine mammals. Geraci [51], published shortly after the Alaska spill, had noted that 15–20 species of marine mammals were at risk in the spill-affected region and that a number of animals had stranded in the first year of the spill response; however, at that time, no pathological or forensic results were available to determine whether the spill had played a role in the observed mortalities.
Loughlin [58] was a focused examination of the effects of the Exxon Valdez on five species of the marine mammal species in Prince William Sound and along the Alaska Peninsula, along with more general results from cetacean surveys and analyses. These were, as the publication date suggests, relatively short-term observations of spill impact, with many results based on gross metrics of mortality with few pre-spill data for comparison. Long-term trends would emerge from research sponsored by the Exxon Valdez Oil Spill Trustee Council (EVOSTC) and other monitoring efforts; ultimately, marine mammals would be among the most impacted of the affected living resources, in terms of the length and severity of effect.

Figure 20.7 Oiled sea otter, Exxon Valdez spill, 1989. Photo: Exxon Valdez Oil Spill Trustee Council.
Sea otters (Enhydra lutris) were the most abundant marine mammal in the spill-affected area and thus were the subject of both short- and long-term studies—as well as a favored media icon for effects of the oil (Fig. 20.7). The EVOSTC documented 871 otter carcasses recovered and estimated that more than a 1000 animals had been oiled in the spill area (another 123 animals died in rehabilitation centers). As many as 40% of the approximately 6500 sea otters resident in western Prince William Sound succumbed to oil exposure.
Bodkin et al. [59] documented that sea otter population in western Prince William Sound had not recovered nearly 20 years after the spill. This assessment was based on demographics surveys, which showed that population numbers on Knight Island—the epicenter of impact for the spill—had not increased despite a steady growth in numbers elsewhere; and on biomarker indications of continued hydrocarbon exposure reflected in elevated levels of cytochrome P4501A (previously discussed as an indicator of exposure in fish). Bodkin et al. [60] continued to track the course of recovery in sea otter populations in Prince William Sound and suggested that a factor impeding that recovery was the presence of pockets of lingering residual oil encountered by the animals when foraging in intertidal and shallow subtidal areas.
It was not until 2013 that the population structure for sea otters in Prince William Sound had achieved a status in terms of numbers, gender, and age that the EVOSTC could declare the affected animals to be recovered—an elapsed time of 24 years. As summarized by the EVOSTC [61]:
Overall, the current population level data for sea otters in PWS are consistent with the EVOSTC definition of recovery for sea otters from the long-term injury incurred in the wake of the 1989 oil spill. The support for this is based primarily on demographic data, including (1) a return to estimated pre-spill abundance of sea otters at northern Knight Island, a heavily oiled area within PWS, and (2) a return to pre-spill mortality patterns based on ages-at-death. Gene transcription rates in 2012 were similar in sea otters from oiled, moderately-oiled and unoiled areas, suggesting abatement of exposure effects in 2012. However, because 2012 gene transcription rates generally were low for sea otters from all areas relative to 2008, these observations cannot be fully interpreted without data from a wider panel of genes. This slight uncertainty with respect to the data from the biochemical indicator is outweighed by the strength of the data for the demographic indicators. The return to pre-spill numbers and mortality patterns suggests a gradual dissipation of exposure to lingering oil over the past two decades, to the point where continuing exposure is no longer of biological significance to the PWS sea otter population.
20.6. The Exxon Valdez and Killer Whales: Circumstantial but Compelling
Orca, or killer whale (Orcinus orca), populations in Prince William Sound represent a rarity in the world of oil spill impact assessment (especially for marine mammals), in that robust pre-spill monitoring information was available. Photographs of individual animals had been collected since the 1970s, and by 1984 there was sufficient information available to define pods that frequented the area [62]; demographic data were collected for 6 pods totaling 122 whales. In the period immediately following the Exxon Valdez spill, researchers focused attention on two groups of whales: the resident AB pod, and the transient AT1 group.
Observations and photographs made it clear that orcas did not or could not avoid oil when it was commonly found on the surface of the water in 1989 (see Figs. 20.8 and 20.9). No animals were found stranded during the spill [63], although three carcasses were found in 1990 and one in 1992 [62]. However, the pre-spill demographic data and information from killer whale studies conducted in other areas of the Pacific coast indicated unusual events and circumstances surrounding the Prince William Sound orcas. In September 1988, 6 months before the oil spill, 36 whales were counted in the resident AB pod; 7 days after the spill occurred, only 29 animals were counted. An additional six orcas went missing between September 1989 and June 1990. For the transient AT1 group (documented at 22 individuals in 1988), the numbers reflected serious instability: Matkin et al. [62] referred to the lack of sightings for any of the nine missing members (one confirmed dead), and expressed doubts that any of the missing whales remained alive. The EVOSTC updated this status and currently lists 15 missing whales, with five carcasses found on beaches [64].

Matkin et al. [65] revisited the status of AB pod and the AT1 group 16 years after the oil spill. AB had not recovered in numbers to its pre-spill level, and the rate of population increase was significantly less than other resident pods. The transient AT1 group continued to decline and in fact was listed as “depleted” under the Marine Mammal Protection Act. Fig. 20.10 [65] illustrates the steep population initial declines for the two groups of Prince William Sound killer whales coinciding with the occurrence of the spill, and shows the long-term trends of recovery (or lack thereof).
It has not been possible to directly link Exxon Valdez oil exposure to the decline and slow recovery of the orcas in Prince William Sound. However, the solid photographic evidence of the whales in oil, and the steep population declines and slow, if any, recovery of the well-monitored AB pod and AT1 group, provide a powerful circumstantial argument connecting the two events. While the recovery status of AB pod is listed by the EVOSTC as “recovering,” that for AT1 group is “not recovering” due to the lack of breeding females and no recent recruitment into the group.

Figure 20.9 Aerial photograph of orca whale (lower center of image) passing through surface oil during Exxon Valdez, 1989. Photo: North Gulf Oceanic Society.

Figure 20.10 Number of killer whales in AB Pod and AT1 group 1984 to 2005 (vertical line: Exxon Valdez oil spill). Reproduced from Matkin CO, Saulitis EL, Ellis GM, Olesiuk P, Rice SD. Ongoing population-level impacts on killer whales Orcinus orca following the ‘Exxon Valdez’ oil spill in Prince William Sound, Alaska. Marine Ecology Progress Series 2008;356:269–81.
Smultea and Würsig [66] used another large oil spill, the June 1990, Mega Borg tanker spill off the coast of Galveston, Texas, to observe behavioral reactions of nine groups of bottlenose dolphins (T. truncatus) in the vicinity of the release. For the purposes of the study, they defined three surface oiling categories: sheen, slick, and mousse (emulsified oil).
Their results were both consistent with and contrary to the previously summarized findings of Smith et al. [53] and Geraci [52], who found that captive bottlenose dolphins were able to detect oil and subsequently avoided surface slicks. That is, Smultea and Würsig concluded from their aerial observations of dolphins near surface oil that the animals could detect both slick and mousse oil on the surface, but did not react to sheens; and perhaps more importantly, they did not necessarily avoid the slick oil that they did detect (they did, however, consistently avoid mousse). Smultea and Würsig also observed behavioral changes in the dolphin groups in the presence of oil, including breakdowns in group integrity, decreased separation between individuals, decreased respiration rates, increased dive times, and increased time to reorient groups.
The Mega Borg spill occurred a little over a year after the Exxon Valdez spill. However, the shoreline cleanup for the latter was ongoing, and with the resource impacts still being tallied, comparisons between the two incidents were inevitable. Smultea and Würsig noted that Exxon Valdez studies of oil-exposed sea otters indicated high incidences of emphysema, petroleum hydrocarbon toxicosis, abortions, and stillbirths. They inferred, from their observations and from previous research, that the greatest threat to cetaceans from the Mega Borg oil was inhalation of toxic fumes and the consequences of that exposure.
20.7. Dolphins and the Deepwater Horizon: “Uncommon Disease Conditions”
It was against this background of limited empirical information and compelling circumstantial evidence of severe oil impacts to cetaceans that the Deepwater Horizon oil spill occurred and numerous cetaceans were observed in oil. The areal extent of oil on the waters of the Gulf of Mexico suggested a likely potential exposure to marine mammals with mostly unknown impact.
Although the initial observations of cetaceans in oil were in offshore waters, the oil eventually moved into nearshore and coastal areas. In portions of Louisiana, heavy amounts stranded in estuaries and marsh habitats, potentially affecting stocks of marine mammals indigenous to those areas. Common bottlenose dolphins, T. truncatus, were observed in heavily oiled areas with oil sticking to their bodies and in apparently impaired health (Fig. 20.11). Clearly, these animals did not/could not avoid the oil.
Barataria Bay was a particularly impacted coastal embayment located in southeastern Louisiana. In addition to receiving a large amount of Deepwater Horizon oil, Barataria Bay was also the site of its own event in July 2010 when an abandoned wellhead was struck and opened by a tug towing a barge. A local resident stock of bottlenose dolphins inhabits Barataria Bay, and thus encountered oil on a sustained basis. Besides obviously being oiled, Barataria Bay local resident dolphins showed overt signs of health issues and thus became the subject of study by a multidisciplinary team of scientists whose results were reported in Schwacke et al. [67].

Figure 20.11 Oiled bottlenose dolphin (Tursiops truncatus), Bay Jimmy, Louisiana, August 5, 2010. Photo: Mandy C. Tumlin/Louisiana Department of Wildlife and Fisheries.
Schwacke et al. compared metrics of health for the Barataria Bay residents to a stock in Sarasota Bay, Florida, where no oil had been observed. This stock was also the most studied dolphin stock in the Gulf of Mexico region. Dolphins in both areas were captured (32 in Barataria, 27 in Sarasota), given veterinary exams, and released. The suite of diagnostic parameters included:
• physical examination
• pulmonary assessment
• hematology
• serum chemistry
• adrenal hormones
• thyroid hormones
• reproductive hormones
• blubber chemistry
The results of the diagnoses showed distinct differences between the two stocks of dolphins, with the Barataria Bay residents characterized by overall poor health condition and afflicted with what Schwacke et al. described as “relatively uncommon disease conditions.” Notable among these was abnormally low levels of adrenal hormones, consistent with a condition called hypoadrenocorticism. Discussing this syndrome in dogs, the Merck Veterinary Manual [68] notes that, “many of the functional disturbances of chronic adrenal insufficiency are not highly specific; they include recurrent episodes of gastroenteritis, a slowly progressive loss of body condition, and failure to respond appropriately to stress.” Hypoglycemia (low blood sugar) and hyperkalemia (elevated blood potassium) were also observed in the Barataria Bay dolphins, and are conditions consistent with compromised adrenal performance.
Schwacke et al. also found indications of impaired lung performance (i.e., five times more likely to have moderate to severe lung scores compared to the reference Sarasota Bay stock) in the Barataria Bay dolphins, a condition which would be consistent with exposure to petroleum hydrocarbons in other species. In particular, pneumonitis, airway inflammation, and pulmonary edema can occur following inhalation exposure. The lung diseases found in the dolphins from the oiled embayment were, according to the authors, “consistent with exposure to oil resulting in either primary lung injury and secondary pneumonia or primary aspiration pneumonia.”
A subsequent study by Lane et al. [69] examined potential reproductive impacts of oil exposure in the same Barataria Bay stock of bottlenose dolphins. The investigation was based on surveys conducted from vessels between 2011 and 2015. Reproductive status of individuals examined during capture-and-release examinations was determined by ultrasound, and calf survival was evaluated through photo-identification of expectant mothers.

Figure 20.12 Dolphin Y01 pushes a dead calf in Barataria Bay, March 10, 2013. The animal was judged to have suffered two consecutive failed pregnancies. Photo: Louisiana Department of Wildlife and Fisheries.
In the 2011 examinations, 10 female dolphins were determined to be pregnant. One of the animals vanished within six months of being examined and was assumed to have died. Among the others, the reproductive success rate was calculated to be 20%; this was much lower than the rate for the reference Sarasota Bay stock (83%). One Barataria Bay individual, Y01, is pictured below in Fig. 20.12 pushing a neonate carcass. This was one of the two documented fetal losses, another being a fetal death observed by ultrasound examination. The nature of the losses for six other pregnancies could not be determined; one was the individual mentioned above that disappeared, while the other five mothers were observed after their due dates with no calves and assumed to be failed pregnancies.
Lane et al. noted that oil could affect reproductive success in both direct and indirect ways. They cited other mammalian studies in which oil exposure was linked to maternal, fetal, and neonatal loss, but also suggested that the impaired overall health diagnostics documented by Schwacke et al. could have adversely affected reproductive success. Bacterial infection by Brucella sp. was additionally cited as a potential dolphin pathogen that can cause abortions, stillbirths, and weak calves. Although there was no forensic evidence of elevated incidence of Brucella in Barataria Bay, they pointed out that reproductive impacts could occur in infected animals without clinical evidence of disease. The implication was that immune system impairment from oil exposure could have increased susceptibility and infection in the dolphins, resulting in the impacts to reproductive success.
Substantially higher than normal dolphin mortalities in Louisiana, Mississippi, and Alabama between 2010 and 2014 led NOAA to declare an “Unusual Mortality Event” (UME) for cetaceans (defined under the Marine Mammal Protection Act as “…a stranding that is unexpected; involves a significant die-off of any marine mammal population; and demands immediate response”). As noted by Venn-Watson et al. [70], disease incidences such as brevetoxicosis (poisoning by red tide dinoflagellates) or morbillivirus (viral infection causing measles in humans, and lung, brain, and immune system impacts in cetaceans) had been confirmed or suspected as causal agents in most (82%) of previous dolphin mortality events in the Gulf of Mexico region. It was therefore important to evaluate whether any of these were factors in dolphin deaths observed in the Deepwater Horizon time frame. However, Venn-Watson et al. determined that Brucella and morbillivirus infections were relatively low in dolphins found dead in Louisiana, Mississippi, and Alabama (7% and 11%, respectively), and biotoxins of algal or planktonic origin were low or below detection. These levels permitted brucellosis, morbillivirus, and biotoxins to be ruled out as primary causes of death in the stranded animals.
Animals in the defined UME area did have a higher incidence of bacterial pneumonia relative to animals from outside the UME area (22% vs. 2%). In UME dolphins diagnosed with pneumonia, 70% were afflicted to the extent that it was determined to have caused or contributed significantly to death. The UME dolphins were also found to have thin adrenal cortices (33% vs. 7%). The adrenal cortex mediates a number of critical processes in an animal, including stress and immune responses, glucose metabolism, and sex hormones.
The health assessment of northern Gulf of Mexico coastal bottlenose dolphins was complicated (as oil spill impact studies inevitably seem to be) by the co-occurrence of other independent physical stressors: a period of sustained cold weather along the Gulf of Mexico coast in the first months of 2010; and an unusual cold freshwater input, or freshet, into the Mobile Bay watershed in early 2011. A record mortality of adult dolphins (as well as finfish, sea turtles, shorebirds, and manatees) occurred in the Gulf of Mexico just before the Deepwater Horizon spill, continued through the active spill period, and in the first reproductive season following the spill, a very high number of dead perinatal (<115 cm) dolphin calves—6 times the historical average—were found primarily on the Mississippi–Alabama coast. Carmichael et al. [71] analyzed whether the cold-water events may have played a role in the spike in deaths of fetal and neonatal bottlenose dolphins in 2011. They could not definitively link the deaths to a single stressor, be it the oil spill or a weather-related event. But they suggested that “strong observational evidence” existed that several stressors played a role in the adverse impacts to a highly stressed group of animals.
Similarly, Lane et al. [69] concluded that while it was not possible to determine whether the reproductive failures they documented were direct or indirect effects of oil exposure, it was not surprising given the other health impacts that had been determined for the Barataria Bay dolphins. The combination of high mortality rates and impaired reproduction, however, was not regarded as a promising formula for long-term success of this group of animals.
This review has shown the progression in knowledge through the last 30 years about oil and cetaceans, and bottlenose dolphins in particular. Whether this progression, which reflects a substantial shift in thinking, constitutes a new paradigm as we defined earlier, remains to be seen; nonetheless, we have moved from a belief, supported by science at the time, that cetaceans could and would avoid oil exposure and its subsequent potential toxicity, to the realization that not only do cetaceans not necessarily avoid oil, they appear to be seriously affected by exposure to oil, with both short-term and long-term consequences at the individual and population levels.
20.8. Oil and Sea Turtles: Known Risk but Little Information
As noted at the beginning of the chapter, the constrained nature of this review of oil effects necessitated a selective discussion of the state of knowledge. With the ongoing burst of research activity initiated by the spill in the Gulf of Mexico, along with the rise in prominence of certain “new” oils (e.g., Alberta oil sands products and Bakken crude oils) due to improved production efficiencies and attendant shifts in global petroleum markets, large amounts of oil-related environmental fate and biological effects information continues to be generated and reported. However, the scale and duration of the spill response in the Gulf of Mexico, with its subsequent NRDA studies, have yielded some insights into oil and spill response effects that are based less on the toxicological science of exposure than on the observational and empirical outcomes of large numbers of resources of interest being exposed for an extended period of time in an actual spill scenario.
The latter has been the case for sea turtles, oil, and the Deepwater Horizon. In the context of oil spill effects, sea turtles share some characteristics with marine mammals. They are “charismatic” and well known to the public; they range throughout portions of the marine environment likely to be impacted by releases of oil; and their protected status and diminished populations have limited research and the available empirical information related to oil effects. However, additional features of sea turtle life history elevate their apparent risk from spills, in that their nesting behavior on sand beaches potentially exposes them to additional shoreline oil and cleanup impacts.
The Deepwater Horizon spill-affected sea turtles and afforded the opportunity to evaluate the state of our knowledge with respect to oil effects on this important group of animals. That they are at risk from oil spills was not a recent epiphany; their risk has been explored, in a largely speculative way, for many years. In 2003, NOAA produced a response-oriented planning guide [72] that consolidated information about features of sea turtle biology and behaviors that intersected with oil spills. Review of oil spill case histories revealed few reported impacts to exposed sea turtles, and a single laboratory effects study [73]. Nevertheless, several key points were identified [74]:
• Sea turtles are highly sensitive to chemical insults such as oil.
• Areas of oil and gas exploration, production, transportation, and processing often overlap with important sea turtle habitats.
• Sea turtles are vulnerable to the effects of oil at all life stages.
• Several aspects of sea turtle biology and behavior place them at particular risk, including lack of avoidance behavior, indiscriminate feeding in convergence zones (areas where two different and distinct bodies of water meet), and large predive inhalations.
• Known oil effects include egg mortality and developmental defects, direct mortality, and negative impacts to skin, blood, digestive and immune systems, and salt glands.
When the Deepwater Horizon blowout occurred in April 2010, aerial surveys of the spill area confirmed the presence and oil exposure of sea turtles (Fig. 20.13). Five of six total sea turtle species (loggerhead, Caretta caretta; Kemp's ridley, Lepidochelys kempi; green turtle, Chelonia mydas; hawksbill, Eretmochelys imbricate; and leatherback, Dermochelys coriacea) are known to occur in the Gulf of Mexico, and all are listed under the US Endangered Species Act. The early aerial surveys showed little ability or tendency of any sea turtles to avoid the oil—to the extent that the sheer scale of the release rendered that option remotely possible.
The association of sea turtles with oceanographic convergence zones became a particular concern during the Deepwater Horizon response for several reasons that had been anticipated in the 2003 NOAA oil and sea turtle response guidance, and in the other earlier literature [75,76]: these oceanic features naturally aggregate surface oil and anything else floating on the surface, but are also locations with higher densities of sea turtles, especially earlier life stages. The associations among turtles, oceanic fronts, and floating beds of seaweed had been well accepted, but largely anecdotal. Witherington et al. [77] conducted a more systematic assessment that confirmed and quantified some of the relationships, and reinforced the importance of floating beds of Sargassum seaweed as habitat for young sea turtles.

Figure 20.13 Aerial photo of sea turtle in emulsified oil in the Gulf of Mexico, May 18, 2010. Photo: Brett Dodson/NOAA.
Response-related considerations amplified concerns about sea turtles, in that convergence zones also became targets for oil cleanup activities such as skimming operations, application of chemical dispersants, and in situ burning of oil at sea. This made sense: natural aggregations of oil were the ideal places to concentrate recovery and remediation efforts. But oil, chemical dispersants, and burning fouled the seagrass beds that were critical habitat for sea turtles, and the continued use of their favored habitat in the face of the massive efforts to remove oil from the surface of the water was a terrible coincidence. Thus, oceanography, biology, and remediation activity literally converged into the oceanographic convergence zones to place sea turtles at risk, directly and indirectly. As this became apparent to resource trustees and then to the operational response managers, aerial overflights were coordinated with on-water vessel reconnaissance to: first, locate convergence lines; and second, locate turtles in the aggregated oil contained in the convergence zones (Figs. 20.14 and 20.15).
A total of 574 live sea turtles were recovered at sea, primarily during the first 3 months of the spill. Over half (317) were Kemp's ridley, and another 17% (220) were green. The remaining 37 recovered turtles were about evenly split between loggerhead and hawksbill. Around 80% (464) of the turtles recovered showed some degree of external oiling [78]. Beside the potential effects of the direct oiling, turtles were also subjected to secondary thermal impacts from absorption of solar heat into the dark masses of oil (Fig. 20.16) that surrounded turtles, as well as reduction in the quality of their favored Sargassum bed habitat. Oiled animals were transported to shoreside cleaning and rehabilitation facilities (Fig. 20.17) in Louisiana and Mississippi where they were externally and internally cleared of oil and stabilized before ultimately being released back into the Gulf of Mexico.
Experience from prior spills and results from the single laboratory study that exposed sea turtles to oil (i.e., Lutcavage et al. [73]) had suggested that external/dermal sensitivity to contact with the chemicals in oil, as well as potential organ toxicity, would be problematic during the Deepwater Horizon—especially with the prolonged and frequently heavy exposures encountered in the convergence zones. However, during the peak of the sea turtle recovery and rehabilitation effort, these impacts were not observed. Moreover, the turtles that were brought into the centers for cleaning and rehabilitation were surprisingly hardy, and responded well, with low mortality and rapid recovery. Indirect thermal, and fouled habitat impacts of oil in Sargassum beds, were judged to be of greater concern for the welfare of the sea turtles in the oil-fouled region.

Figure 20.14 Surveying convergence line for oiled sea turtles, Gulf of Mexico. Photo: Blair Witherington, Florida Fish and Wildlife Conservation Commission.
Ultimately, the most significant development affecting the health of sea turtles was the securing of the wellhead release on July 15. This quickly reduced the surface expression of oil in the northern Gulf of Mexico, and resulted in much improved convergence zone/Sargassum bed habitat for the turtles. Within a few weeks, sea turtles with sufficient external oiling to require transport to a rehabilitation center were encountered only infrequently, allowing this phase of the wildlife response to be curtailed.

Figure 20.15 Dr. Brian Stacy, NOAA veterinarian, with sea turtle recovered from convergence zone in the Gulf of Mexico. Photo: Blair Witherington, Florida Fish and Wildlife Conservation Commission.

Figure 20.16 Infrared thermometer showing surface temperature (127.2°F) of surface oil in the Gulf of Mexico during the 2010 Deepwater Horizon spill. Photo: Brian Stacy, NOAA.
Meanwhile, in addition to the large volumes of Deepwater Horizon oil potentially encountered by sea turtles in the pelagic and coastal marine environments, the cycles of nesting on sand beaches throughout the affected coastline of the Gulf of Mexico were cause for concern for sea turtle managers and the spill response structure. The limited research and previous spill experiences had suggested that sea turtle eggs and hatchlings were especially sensitive to oil exposure and susceptible to the seemingly inevitable beached oil. The continuous nature of the oil release between April and July of 2010 increased the potential for large amounts of oil to strand on the beaches of the northern Gulf of Mexico, where hundreds of sea turtles (primarily loggerhead) would nest, presenting the dilemma of remedial versus preemptive actions, i.e., should the nests remain in place and oil dealt with as necessary, or should a plan to physically remove and relocate nests be implemented?

Figure 20.17 Sea turtle veterinary care administered at the Audubon Nature Institute in New Orleans, August 2010. Photo: Gary Shigenaka, NOAA.
The option of nest relocation was portrayed in the 2003 NOAA guidance document on oil and sea turtles [72] as a spill response measure of last resort. Perhaps as a measure of how serious the potential for widespread shoreline oiling was judged to be in 2010, nest relocation was the action chosen for 278 sea turtle nests from Alabama and Florida Panhandle beaches, representing 28,568 eggs. Eggs were excavated from nests (Fig. 20.18), packed into styrofoam crates, and trucked in temperature-controlled containers to the Kennedy Space Center, where they were incubated and hatched. The hatching success/release was a robust 14,676, or 51%. Despite these impressive metrics: the larger questions of benefit to the Gulf of Mexico stocks of sea turtles, or even where the relocated turtles will return when they are ready to reproduce, remain unknown and may never be resolved. It will be at least three decades before any of the Deepwater Horizon year class sea turtles mature and return to lay eggs; the hatchlings were not marked or tagged. Because of the deep uncertainties surrounding the viability of these animals, the Deepwater Horizon NRDA simply considered the relocated eggs as resources lost to the spill.
The Deepwater Horizon sea turtle experience was remarkably informative, but as the preceding narrative reflects, it was not anchored in the toxicology or the mechanistic pathobiology that characterized the stories of fish embryo and marine mammal effects that preceded it in this chapter. At this writing, the sea turtle story is more of a forensic investigation that was built on older research and data either validated (importance of and amplification of risk represented by oiled convergence zones)—or not (assumed sensitivity of sea turtles to dermal oil exposure). Laboratory results from projects extrapolating insights from surrogates such as terrestrial turtle species, novel new techniques that might define impact at lower levels of organization than the organism, or correlation studies (along the lines of the long-term Exxon Valdez orca population monitoring surveys) may provide supporting or confounding results to inform the oil and sea turtle story. But given the theme of this chapter: in 10 years, we may well be surprised, but smarter.

20.9. Conclusion
In the preceding sections, you have read about three different approaches to understanding the effects of oil. The fish studies are largely based on controlled laboratory studies to determine mechanisms of effect at the suborganismal and even subcellular level. Exposures to PAHs are (mostly) well controlled and measured, and approximate those that might be encountered in the real world, during an actual spill. The effects noted would clearly compromise the health of individuals, and presumably of communities and populations. However, the challenge remains to show how the effects so well documented in the laboratory translate to populations and ecosystems.
The marine mammal studies, beginning with the Prince William Sound orca surveys, began at the opposite end of the scale of biological organization. That is, instead of subcellular and molecular effects, the apparent effects of exposure to an oil spill manifested themselves in aberrant and unprecedented changes in normally stable population levels of well-monitored stocks of whales. There were, however, no dead orcas to study in order to provide a more definitive link to oil exposure as a potential cause for population declines. The situation in Barataria Bay with bottlenose dolphins, however, was different. Researchers had evidence of individual illnesses and deaths in an established stock, and were able to sample both living and dead animals. This permitted the creation of a toxicity construct consistent with veterinary pathology and with the observations from the field. It represents a more complete story, but one that can be augmented and strengthened over time.
Finally, the sea turtle experience was almost completely derived from field observations and recoveries, with some postmortem necropsy results and data derived from living animals brought in for rehabilitation. A large amount of information was generated on effects of oil exposure to individuals, though much less so on potential population effects. The net effect of relocating nests and release areas for a considerable number of loggerheads may well never be determined. Nevertheless, the spill represented an opportunity to validate some of the speculative assumptions we had made for oil spill response; some were supported, others were not.
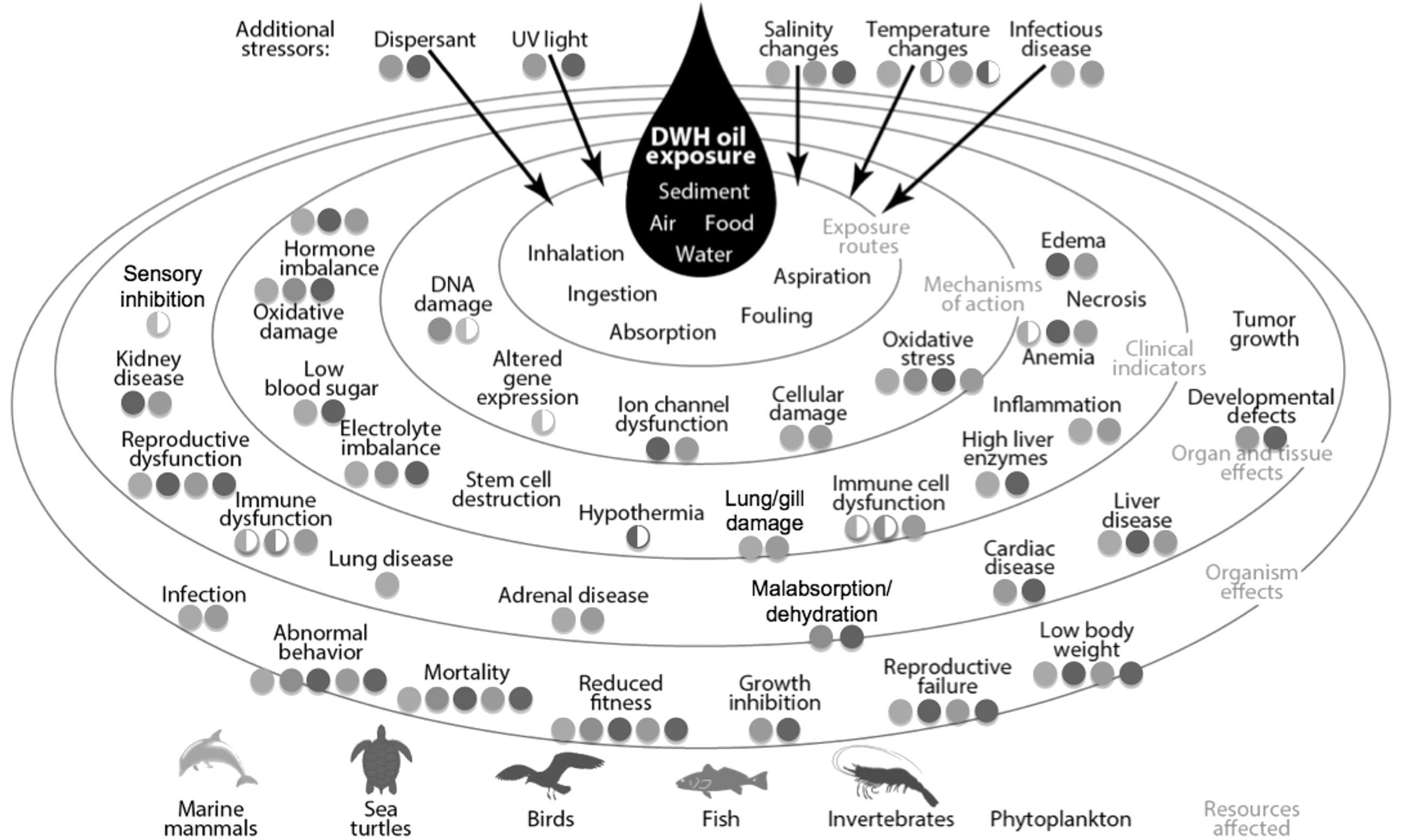
The approaches to these three taxa have been very different. The results and demonstrated or inferred mechanisms of toxicity are distinct as well. During oil spill response, biologists typically counsel against extrapolating effects in one organism or population to another, or from one oil type to another. There is a wealth of information in the scientific literature and in the gray literature to support the existence of differences in oil sensitivities in different organisms, and differences in toxicities of different products.
Yet, those who have been able to view the universe of effects studies from the Deepwater Horizon NRDA, which span many more taxa and levels of biological organization than discussed in this chapter, suggest that there are patterns of commonality in the vast amount of research conducted on behalf of that effort. This “constellation of relationships” (as they termed it) is portrayed as Fig. 20.19.
The graphic conveys the notion that despite differences in oil effects in different organisms, the opportunity to study many different resources affected by the same oil spill under a common research umbrella has suggested that similarities exist in terms of how oil exposure harms dissimilar taxa. While this is not exactly the oil impact equivalent of the Grand Unification Theory in particle physics—it may not even rise to the level of a paradigm shift for how we consider oil toxicity—it may be one step closer to a more consistent model of oil effects. We all benefit from understanding the extent to which we can generalize about oil impacts—and just as importantly, the reasons why we cannot.
..................Content has been hidden....................
You can't read the all page of ebook, please click here login for view all page.