
Chapter 5
Microbial Cell Refining for Biomass Conversion
Abstract
Cell refining can use renewable biomass resources as raw materials and decompose biomass through complex microbial metabolic networks and regulatory networks to synthesize a series of products to replace fossil energy and chemical products. The efficient and directed production of target compounds by microbial cells during a biorefinery requires the interference and changing of the original microbial control system. In essence, the core of cell refining is strains selection, which includes the creation of a new metabolic pathway, which enables the construction of the metabolic network model, and allows the design of new products by altering or reducing the metabolic pathways. Therefore, on the basis of the regulatory mechanism of microbial metabolism, achieving both a high production yield and a high production intensity of target metabolites by directed changing and optimization of physiological function of microbial cells have a significant effect on the biomass conversion technologies using cell refining as the core content.
Keywords
cell refining
systems biology
artificial cell
intercellular synergy
cell refining plant
5.1. Biorefinery
In 1982, the concept of biorefinery was first proposed in Science, in which agricultural waste, plant-based starch, and lignocellulosic materials were used as raw materials to produce chemicals, fuels, and bio-based materials. Biorefinery was defined by US National Renewable Energy Laboratory as something using biomass as feedstock, combining biomass conversion processes and equipment comprehensively to produce fuel, electric energy, and chemical products. Due to the current crisis of energy, resources, and worsening environmental issues, the concept of a biorefinery has become a strategic research around the world, leading to the launch of national biorefinery plans in many countries (Kamm, Gruber, & Kamm, 2007).
Microbial cell factories are the core of biorefinery technologies and the platform and material basis of cell refining. Cell refining is the process during which renewable resources were used as raw materials for energy, drug, and chemical materials production for human by microbial cell transformations. The first electron microscope made people understand that the eukaryotic cell is a complex open system consisted by cell film and passages, whose internal structure is similar to a factory. In the biological world, the specific surface area, transforming ability, reproduction rate, and other indicators of microorganisms, exceeded above all creatures, and thus microorganisms have a strong self-regulation and environmental adaptability. Based on these characteristics, “microbial cell factories” successfully realize the cell refining which serves human needs, such as Actinomycetes. sp. (antibiotics production), Penicillium. sp. (penicillin production), Corynebacterium glutamicum (lysine and glutamic acid production), and Aspergillus niger (citric acid production). In order to exploit microbes’ potential more efficiently, scholars screen finer strains to produce better products by utilized known metabolic regulation mechanism to transform various microorganisms based on well-known metabolic mechanism. The process is shown in Fig. 5.1 (http://news.sciencenet.cn/sbhtmlnews/2010/5/232185.html).
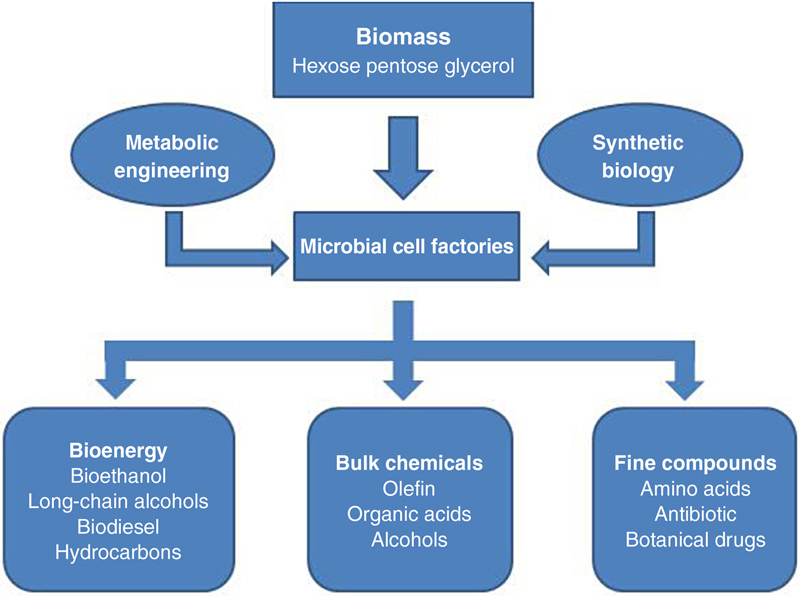
Figure 5.1 Schematic diagram of the cell refining process.
Future biochemical conversion of biomass is a technology combination of bioconversion and chemical cleavage, including improved pretreatment and fractionation of lignocelluloses, optimized design of reactors for conversion of renewable feedstock, and improved biological catalysts and catalytic processes. Cell refining is a typical cross-domain of interdisciplinary research and its development needs joint efforts of experts from biology, chemistry, and engineering disciplines and other areas of technology, which integrated the refining process effectively.
Up to 200 billion tons C are fixed annually on the earth through photosynthesis of plants, with the energy of 3 × 1018 kJ, about 10 times the energy consumed worldwide per year. Among them, straw is the largest biomass resources efficiently collected. More than 700 million tons straw were collected in China per year, which is expected to be conversed into raw material of bio-based chemical products to solve resources and energy crisis currently in China. However, although scientists around the world have studied this aspect for many years, the high-value utilization of straw resources has still been not achieved. The main obstacle is the technical and economic problems of straw conversion, which is a world scientific problem and engineering challenge.
Sugar and starch were the main raw materials in traditional fermentation industry. The current rise of straw and other biomass refining is aimed to substitute the glucose starch with the glucose derived from cellulose. To replace the traditional structure of the petrochemical industry by straw and other biomass industry, the issue to use biomass as the new commonly used feedstock of biological and chemical industry should be broken through. Chen, Li, & Liu (2011) had analyzed the differences of chemical compositions, physical properties, and cellulose hydrolysis of straw from the level of molecule, cell, and tissue, revealing that the nonuniformity of straw is the primary cause of the difficulty of value-added-use of straw resource (Chen & Qiu, 2007; 2010). They have developed traditional straw utilization patterns based on the whole plant into a value-added conversion system with comprehensive biomass utilization by component fractionation and stratified multistage conversion (Jin & Chen, 2006; 2007; Chen, 2010). Based on their systematic reach they indicated the research direction for the high-value utilization of straw resources by providing a sophisticated scientific theory and engineering application of straw component fractionation—stratified multistage conversion for the establishment of a new biomass feedstock refining industry (Chen, 2006, 2008a,b), creating a new eco-industrial model of straw and other biomass refining for biobased products.
5.2. Cell Refining
The production processes of the cell refining platform include input output of organic raw materials, activation of genetic information, formation of a metabolic network, formation of metabolic material, and output of product. The microbial cell factory is the basic platform to achieve the goal of biomass biochemical conversion. However, the enzyme species of the wild-type microorganism in nature do not perform effectively and are not favored by industrial production.
In order to transform microbes into microbial cell factories for biorefining, the latest technologies and methods, including genomics, metabolic engineering, systems biology, and synthetic biology, should be the tool for analysis of the essence of microbial gene, protein, and metabolic processes network, developing a better understanding of the microbial metabolism’s ability from the molecular, cellular, and ecosystem level. Based on this, scholars could redistribute the material and energy flows of microbial cell metabolism through the reorganization and optimization of manual control and fully exploit the microbial capabilities of decomposition, transformation, and chemical synthesis, and effectively prepare bioenergy and alternative petrochemical raw materials. Thus, the abundant biomass resources could truly become a substitute of petroleum industrial raw materials, and efficiently be prepared to bioenergy (biogas, biohydrogen, bioethanol, biodiesel, etc.) and platform chemicals (ethylene, lactic acid, fumaric acid, butyl acid, furfural, 1,3-propanediol).
Metabolism is the basic feature of life phenomena. As a three-dimensional cell factory, the major metabolic pathways of microbes are related directly to the utilization of raw materials and product synthesis. However, the intracellular metabolic pathways are not independent and in complex metabolic network by various regulation. Regulation is a complex process at the transcriptional level and translation level. The existing regulatory system of microorganisms must be adapted, changed, or modified to improve the efficiency of cell factories.
Genetic and environmental factors determine the microbial conversion capabilities. From a genetic perspective, the result of long-term evolution makes metabolic function a benefit for the microbial body. The metabolic processes are in the most economical state and do not excessively accumulate unfavorable metabolites. However, cell refining requires microbial cells to change the original metabolic pathways and accumulate a large number of expected metabolites (secondary metabolites). Thus, in order to achieve the rational regulation of microbial metabolic function, an in-depth understanding of the molecular mechanism of metabolic pathways, the microbial genomes and their interactions with the corresponding enzymes is required.
5.2.1. Metagenomics of Cell Refining
Metagenome, also known as metagenomics or environmental microbial genomes, is the sum of the entire tiny DNA in biological environment (currently mainly bacteria and fungi). Metagenomics aims to study the microbial population genomics in environmental samples without the microbial cultivation stage. During the analysis process, the total DNA in the environment was directly extracted. Genetic screening and sequencing functional analysis were employed to study microbial diversity, population structure, evolutionary relationships, microbial functional activity relationships, and mutual cooperation relations between microbes and the environment. The appearance of metagenomic technology makes people aware of studying uncultured microorganisms, which account for more than 99% of all microorganisms, significantly enhancing the detectable space of microbial genes. Overall, the current application of metagenomic techniques can be divided into two aspects: developing the required function proteins by screening functional genes; and exploring the interactions between microorganisms and the surrounding environments through the analysis of metagenomic library to understand the microbial world more objectively and comprehensively.
5.2.1.1. Basic Strategy of Metagenomics
The basic strategy of metagenomics is to:
1. extract the total DNA or mRNA from the environmental samples as completely as possible;
2. use appropriate vectors to clone the genomic DNA into the mode microorganisms to establish a macro genomic DNA library or a cDNA library; and
3. give new traits to host cells by exogenous gene or screen based on some known DNA sequence to find a new target gene clusters or biologically active substances.
Based on the general process of screening new genetic resources and expressed products from Rumen microbes, the construction policies of metagenomic library are summarized in Fig. 5.2 (Wang, An, & Liu, 2010).
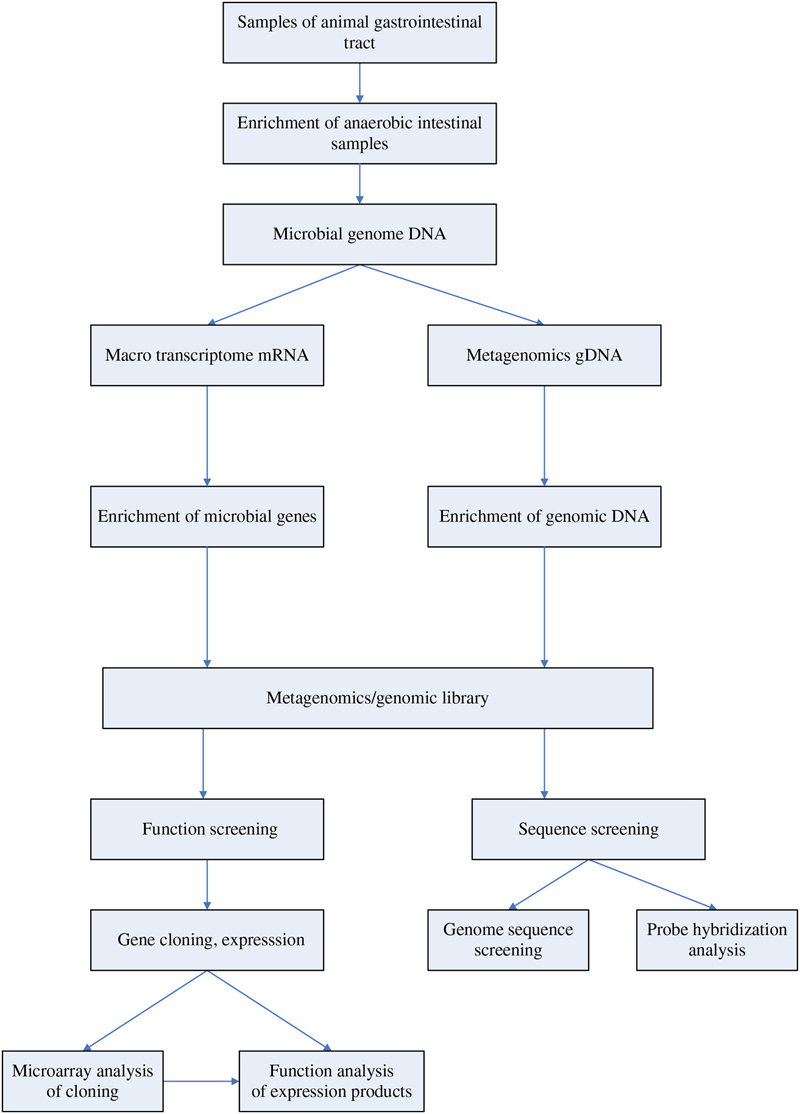
Figure 5.2 Construction and screening process of animal gastrointestinal metagenomic library.
Metagenomics analysis will enable us to deeply understand the diversity of rumen microorganisms, population structure in the system, evolutionary relationships, functional activity, and mutual cooperation relations, and to build control means of multiscale anaerobic digestion process based on reactor operation, changes of active bacteria, and functional changes in gene expression, providing a guiding role for the anaerobic microbial screening and biomass anaerobic conversion process control.
5.2.1.2. Construction of a Metagenomic Library
Construction of a metagenomic library follows the basic principles and techniques of molecular cloning. Some special purpose steps and strategies should be taken according to the specific characteristics of the environment samples and the goals for database building. It is one of the key processes for the construction of metagenomic library to obtain the high purity, high molecular weight, and high concentrations of total DNA from environmental samples, which needs to extract the total DNA from the samples entirely so as to maintain its large fragments of DNA and to obtain the complete target gene or gene cluster.
Currently the main methods used for DNA separation and extraction from environmental samples, such as soil are the situ lysis and ectopic lysis methods. Environmental samples in the situ lysis method (also known as the direct lysis method) were suspended in lysis buffer directly for extraction and purification, which is suitable for plasmid or phage as cloning vectors. This method has the advantages of easy operation, low cost, and high DNA extraction rate, while a smaller fragment of DNA is extracted (1∼50 kb) due to the strong mechanical shear. Ectopic adhesion lysis isolates microbial cells in samples of soil and sediment first, followed by a more moderate approach to extract DNA, known as the microbial cell extraction method. For the vector, the construction of a metagenomic library currently uses a plasmid, cosmid vector, and bacterial artificial chromosome. A bacterial artificial chromosome vector can clone large fragments of DNA, increasing the chance for complete guidance molecules, such as genes or gene cluster encoding antibiotic synthesis pathway, which shows a considerable advantage in the cloning and expression of macrogenome.
To improve the expression level of the gene so as to facilitate the activity detection of recombinant clone, the expression vector was employed by some researchers to construct a metagenomic library directly. In addition, the shuttle vector can expand the host range in favor of expression of exogenous genes. Screening factors of the transformation efficiency, the stability of the recombinant vector and the expression of macro gene and target traits should be taken into consideration in selecting a host strain (e.g., antibacterial activity). The results showed that the active substances produced by different types of microbial species are significantly different. Therefore, the target host strain should be chosen according to the purpose, for example, 70% of the antibiotics from actinomycetes, Streptomyces sp., should be selected as a host bacteria to look for antibacterial, antitumor active substances while E. coli is used for the new enzyme screening.
5.2.1.3. Screening of a Metagenomic Library
Currently metagenomic library screening methods are divided into sequence-based screening, function-based screening, compound structure screening, and substrate-induced gene expression screening.
Screening based on the sequence is the process of designing a probe or PCR primers according to the sequence of associated functional genes to obtain a target gene from the gene library. Compound structure screening is the process of screening target genes by comparing the chromatogram of fermentation broth or extracting solutions from host cell with/without transferred exogenous gene. However, the substances screened by this method may not have the activity. Substrate-induced gene expression is the screen process using substance-induced catabolic gene clones, which can be used for the screening of enzyme activity.
Metagenomics technology fills the blank of uncultured microbial research and has become a focus of international microorganism study, forming a new path to find new genes, develop new biologically active substances, and study the diversity of microbial communities (Zehr, Bench, & Carter, 2008; Mori, Mizuta, & Suenaga, 2008). Metagenomics technique does not rely on training while extracting genomic genetic material directly from the natural environment microorganisms to analysis the microbial population, which boosts better awareness of the knowledge of microbial population ecology and evolution, maximizes the excavation of microbial resources. The technology makes the study of environmental microbes focus not only on “determining what they are,” but also on “determining what they can do” without the training process (Wexler, Bond, & Richardson, 2005; Xu, Duan, & Zhou, 2006).
Of course, the study of metagenomics faces many challenges. A metagenomic library consists of uncultured microbial genetic information, rather than the bacteria itself. The genetic background of these microorganisms is unknown and unable to study regulation of gene expression. It is possible that an exogenous gene is not expressed or undetectably expressed in the host cell. Thus, the metagenomic library screening may miss many natural products of environmental microbes, which will affect the results of the metagenomic study. However, with the improvement of metagenomic technology, progress will continue to enhance its application value.
5.2.2. Metabolic Engineering of Cell Refining
The advent of genetic engineering has greatly promoted the development of microbial fermentation industry, which makes genetic modification of specific enzyme reactions in the microbial metabolic pathway possible. However, the microbial fermentation involves the synergistic action of multiple enzymatic reactions in the metabolic network of microorganisms. Genetic transformation of a single (or multiple) enzymatic reaction is not sufficient to improve fermentation performance. Extremely rich biodiversity of microorganisms determines their metabolites with diversity. With the acquirement of sufficient sequence information, metabolic pathways of these organisms will be reconstructed, providing effective strategies for us to develop these creatures. In 1991, Jay Bailey and Gregory Stephanopoulos first proposed a metabolic engineering concept: “using recombinant DNA technology to manipulate enzymes, transport, and regulatory functions, thereby to improve the cell activity.” With the development of technology, the definition of metabolic engineering has been corrected continually by later scholars.
The essence of metabolic engineering is the quantitative analysis and control of the metabolic flux for the metabolic reconstruction to maximize the yield of the desired metabolites. Unlike traditional mutation breeding techniques, it is a purposeful and rational reconstruction, involving physiology, molecular biology, biochemistry, and biological pathway engineering and other disciplines.
The main contents of metabolic engineering theory involves:
1. biosynthesis and metabolism network theory;
2. metabolic flux analysis, nodal analysis;
3. carbon resources, respiratory system, redox redesign; and
4. action mechanism of center metabolism and related metabolic analysis.
It is worth noting that the two differences of metabolic engineering compared with genetic engineering are as follows.
First, metabolic engineering is the systematic study of cell metabolism networks, putting more emphasis on the “integration” role of multiple enzymatic reactions. Second, after the completion of genetically engineered metabolic pathways, metabolic engineering should analyze the physiological changes in cell metabolism fluxes in detail to decide the next target for genetic transformation and constantly improve the physiological properties of cells (fermentation capacity) through multiple cycles.
The completion of more and more biological gene sequencing and the progress of functional genomics research introduce the metabolic engineering into a new era. However, how to regulate a cell metabolism network effectively to improve cell performance is always the core of metabolic engineering and the microorganisms’ cell refining.
Principles and methods of metabolic engineering based on the systematic analysis of metabolic networks is to transform the cell metabolism system using recombinant DNA technology, which includes three key steps: first, genetic modifications: construction of recombinant bacteria for improved performance; second, metabolic analysis: analysis of recombinant bacteria metabolic pathways, especially comparing its performance with the original strain performance; third, the design strategy: the design of genetic engineering goal for the next step. These three steps constitute one cycle, which is shown in Fig. 5.3 (Li, 2009).

Figure 5.3 Principles and methods of metabolic engineering.
Metabolic engineering research usually consists of two parts; one is the analysis for cell systems. A cell metabolic network is a very complex system where the metabolism of cells can affect gene expression. In contrast, the expression of the gene controls the associated enzymes in the metabolic pathway and influences the choice of metabolic flux. Therefore, the analysis of metabolic engineering includes not only the intracellular metabolic flux, but also the intracellular gene expression by using transcriptomics and proteomics tools, which could reveal the correlation of gene expression and metabolic pathways. Another part is to genetically transform of engineered strains, change the metabolic flow, open up new metabolic pathways, and build superior strains by the results of the analysis (Zhang, 2009).
5.2.2.1. Metabolic Network Theory
The theory of metabolic networks takes the biochemical reactions of cells as a whole network. The network of the cell metabolism consists of reaction system catalyzed by thousands of enzyme, membrane transport systems, and signal transmission systems with fine adjustment and coordination. The metabolite of shunt metabolic network is called a node and a small number of nodes that play a decisive role for the end products are called main nodes.
5.2.2.2. Metabolic Flux and Intermediate Products Analysis
Metabolic flux analysis is an important means of metabolic analysis. It is assumed that the material and the energy inside the cell are in a steady state. The applications of extracellular substances concentration determination, radiolabeling, and isotopic tracing make the metabolic flux analysis easier.
Through the flow analysis of cell under different situations, such as changes of the culture environment, removal of suppression, increase or decrease of enzyme activity, the node type and the optimal way could be determined. Also through the ways, the results of gene modification and the maximum theoretical yield rate could be estimated. For simple reaction system, satisfactory results could be obtained through accurate analysis and balanced calculation of all metabolic network. However, for more complex metabolic systems, pathways analysis becomes difficult. With the rapid development of computer technology, the methods to analyze the metabolic become diverse and promote a higher degree of automation.
Thus, the flow to the final product from the metabolic intermediates will ultimately determine the yield of the final products. Metabolic flow changes of intermediary metabolites will be inhibited by cells and cause severe interference of intracellular function. Since some intermediate metabolites play an important role in cell signaling and the regulation system, the role of intermediate metabolites in the cell must not be overlooked during the metabolic transformation.
5.2.2.3. Metabolic Control Analysis
Metabolic flux analysis reveals a static distribution of metabolism, and metabolic control analysis aims to the instability of internal and external environment for the cells, revealing the dynamic changes of cellular metabolism. The coefficient of elasticity and flow control are the two main indicators of metabolic control analysis. The elasticity coefficient reveals the effect of metabolic changes on the reaction rates. These two factors are interrelated, and may be measured directly or indirectly (Zhao, Wang, & Chen, 2004). The flow control coefficient is the metabolic flow changes of a branch flow caused by changes of the enzyme amount and is used to measure the control degree of a step enzymatic reaction on the entire reaction system.
On the basis of microbiology and in-depth research of the genome, metabolic engineering techniques study the metabolic pathways of biochemical conversion of specific compounds and develop microbial primary metabolites, secondary metabolites, and important biobased chemicals. In addition, conducting functional studies of biocatalysis, optimizing the strains for essential enzyme protein production, modifying gene of enzyme, and developing new industrial enzymes are all the content of metabolic engineering. Galazka et al. transferred transport protein CDT1 and CDT2 of cellobiose and oligosaccharides from Neurospora crassa into yeast and realized the utilization of cellobiose by yeast. The yeast could take in cellobiose to produce ethanol directly by this modification process, which improved the efficiency of the whole sugar fermentation of cellulose and provided a new idea for large-scale production of biofuels (Galazka, Tian, & Beeson, 2010).
Complex interactions between enzymes, regulators, and metabolites in microbial cells lead to extremely difficult work in optimizing metabolic pathways, which is facing serious challenges in metabolic engineering. The appearance of new methods of genomics, transcriptomics, and metabolomics greatly promotes the optimization of metabolic pathways. In particular, the development of systems biology provides a global scale for a profound understanding of the physiological and metabolic characteristics of microorganisms, creating an unprecedented opportunity for the development of metabolic engineering.
5.2.3. Systems Biology Technology of Cell Refining
Achieving high yields of the target metabolite, a high yield and high intensity of production organic unity based on a better awareness of the regulation mechanism of the microorganism metabolism is the core content for cell refining biochemical biomass conversion technologies and has very important significance.
The cell is a tightly regulated, multichannel, and multilevel network system to a large extent against external disturbance, which undoubtedly reduces the effects of genetically modification. In order to make further progress in metabolic engineering, deepening the understanding of the physiological activity patterns of cells is required. With the wide application of high-throughput experimental techniques, the gradual improvement of bioinformatics methods, and enhanced capacity of excavating valuable information from large data, the understanding of physiological activity law in microbial cells is deepened and the systems biology emerged. This systems biology enables metabolic pathway engineering from the local level to the overall level, and metabolic engineering has thus entered a new stage of development, which is called the system biotechnology era.
Systematic biology is the study of all the components of biological systems (gene, mRNA, protein, etc.), and the subjects of mutual relations between the components under these specific conditions. In other words, the previous experimental biology concerns only with the case of genes and proteins, and systematic biology will have studied all relationships among genes and all proteins (Zhao, Bai, & Li, 2010). Whole research is the biggest feature to use the systems biology approach to study inherent physiological changes in microbes, microbial interactions, and relationships with the external environment interactions between microbes. This method can explore the whole domain of microbial biosynthesis regulatory genes and provide a more comprehensive theoretical basis for species improvement, remodeling and microbial genome expression, and systems regulation.
Traditional methods of strain transformation are random and time-consuming. As in-depth understanding of metabolic pathways and regulatory mechanisms, metabolic engineering achieves the metabolic network optimization by overexpression or knocking down the key metabolic pathway genes. However, it is difficult to achieve the desired results by multiple genes to determine a trait, particularly when there are not suitable target genes to operate for less knowledge about the control mechanism. Development of reverse metabolic engineering is used to analyze metabolic pathway of mutants with improved performance and find the key targets to improve the physiological performance of strain, in order to achieve the targeted reconstruction of a metabolic pathway. This technology also achieves some success, but is still insufficient.
In recent years, cell genome modification and a genome transcription project have provided a new way for bacteria transformation. These directed evolution technologies used in microbial metabolic engineering enable us to regulate gene expression levels more finely, and change the transcriptional level of multiple intracellular genes simultaneously. However, it still involves the nondirectiveness of mutations and the screening of vast mutants. Moreover, due to the interaction and the regulatory mechanisms of metabolic networks inside the cell, some genes which do not relate to the metabolic pathways, the expression level, or expression products may also have a significant impact on the optimization of metabolic networks, while overexpression a single gene or a few genes may cause an imbalance of a metabolic network as a whole, thus affecting the success of metabolic engineering.
To find the key genes affecting metabolic optimized pathways and to achieve the balance of the regulation of metabolic networks are important research goals in metabolic engineering operations. The development of systematic biology provides people with a comprehensive understanding of the cell from the genome-scale metabolic network view, including the structural genes, which are compositions of the metabolic pathway, the complex regulatory mechanisms of cell metabolism, as well as the influence of genetic and environmental perturbations on the overall metabolism of the cells. Systematic biology technology thereby establishes an omics-scale metabolic model, which could evaluate and predict the possible effects of genetic engineering operations and guide the operation methods of metabolic engineering through the metabolic network analysis. Finally, physiological function of cells could be improved, including the yield and efficiency of bioethanol, biobutanol, organic acids, amino acids, and other fermentation products, and obtaining the species processing flow that meets the need of industry.
Typical cellular metabolic engineering strategies in cell refining include the following three steps:
1. Construction of the initial engineered strain. This stage is similar to the traditional metabolic engineering strategy previously mentioned: transforming local metabolic pathways by analyzing the structure of the local metabolic network (such as reducing by-products by knocking out the competition pathway), and optimizing the physiology performance of a cell (such as lifting product toxicity and feedback inhibition effect). The schematic routes of the strain transformation process guided by systematic biology are shown in Fig. 5.4.
2. Systems analysis at genome level and computer simulation of metabolic analysis. As mentioned previously, high-throughput omics analysis techniques can effectively identify the new target genes and pathways that improve the production capacity of cell fermentation. At the same time, by using the metabolic network model at the genomic level, some other new target genes could also be simulated and analyzed. It should be emphasized that the target genes identified by these two systems analysis methods are not related to the local metabolism pathway; the traditional metabolic analysis makes it hard to identify them.
3. Optimization of industrial fermentation processes. The first and second rounds of microbial fermentation are carried out under laboratory conditions, which have many differences in the fermentation performance compared to large-scale industrial fermentation. Scale-up is often associated with the generation of high concentration by-products, and therefore the next round of metabolic engineering to optimize the fermentation ability of bacteria is still necessary.

Figure 5.4 Strain modification process by systems biology.
In recent years, there has been great progress to utilize information acquired from systematic biology research to guide metabolic engineering operations and directional design of a new metabolic pathway for the biochemical conversion of biomass in refining cells.
Succinic acid in the krebs cycle is a platform compound formed by the reduction of oxaloacetate. Hong et al. determined all of the 2,314,078 gene sequences of Mannheimia succiniciproducens survival in rumen with hypoxia and a carbon dioxide-rich environment. They not only defined the genetic map of the bacteria, but also elucidated the main metabolic pathways of bacteria adapted to survive in the rumen. The bacteria can produce large amounts of succinic acid accompanied by the generation of some other organic acids. In order to improve the production of succinic acid, based on the results of the complete genome sequence, a metabolic model containing 373 reactions and 352 metabolites was constructed. The metabolic flux analysis showed that the carbon dioxide and phosphoenolpyruvate carboxylation into oxaloacetate were equally important to cell growth. Based on the results, from the view of the genome, improvement strategies of M. succiniciproducens were proposed (Hong, Kim, & Lee, 2004).
Saccharomyces cerevisiae is the most commonly used industry host for fuel ethanol production and also one of the microbial model organisms. Jens Nilson and his research group (Bro, Regenberg, & Forster, 2006) built a biological model system of S. cerevisiae in genome scale. They designed a metabolism engineering operating strategy to reduce the yield of glycerol and improve the ethanol yield. Further metabolic engineering study showed that overexpression of glyceraldehyde-3-phosphate dehydrogenase gene in S. cerevisiae decreased the glycerol yield by 40% and increased the ethanol yield by 3%. In the mixed sugar fermentation system of engineered strain with overexpression of the gene, the production rate of ethanol increased by 25% with substrate of glucose, and xylose.
5.2.4. Cell Processing and Synthetic Biology
Synthetic biology is evolved from people’s knowledge and understanding of life over a vast length of time, which is based on deepening understanding of the structure and function information about the genes and proteins and other basic constituent elements of life. Synthetic biology research emerged in response to innovative thinking and technology with the material of natural biological elements and support platforms of molecular biology and genetic engineering and other modern biotechnology. In 2000, the first successful artificial manufacture, a circuit similar gene regulatory networks, marked the birth of synthetic biology (Gardner, Cantor, & Collins, 2000; Elowitz & Leibler, 2000). But the real access of synthetic biology to the public was from the research reports of US biologist Craig Venter (Gibson, Glass, & Lartigue, 2010). A mycoplasma genome of 1080 kb was synthesized in the laboratory and then implanted into the goat mycoplasma cell without genetic material, creating the first “artificial single-cell organisms,” which declared that the birth of the first gene does not rely on natural gene template and the synthetic bacteria that had the ability to self-replicate.
Genetic engineering technology is the core of synthetic biology. Based on the techniques of molecular biology, genomics, bioinformatics, and systems biology, synthetic biology designs, transforms, and reconstructs biological molecules, biological components, bioreactor systems, metabolic pathways, and processes, as well as cells with vitality, individuals, and other biological systems, and provides a biological way of manufacturing low-cost biological drugs, chemicals, functional materials, or energy alternatives (Wang, Peng, & Hu, 2011).
Synthetic biology is different to traditional biology, which studies the internal structure of organism by anatomy. Synthetic biology is the study from basic organism elements to the artificial life system. Synthetic biology and genetic engineering are different in the connotation. Genetic engineering is the continuation, transformation, and transfer of the genes from one species to another while the aim of synthetic biology is to assemble the various components to create artificial biological systems and make the components able to run in vivo as the circuit and complete a variety of biological functions (Liu, Du, & Zhao, 2011).
The goal of synthetic biology research is very clear: the synthesis of new life forms, or transformation of existing life forms to achieve new functionality. To achieve these goals, scientists explored from different levels and aspects. Currently, synthetic biology research is focused on three aspects (Zhang, Chang, & Wang, 2010): (1) standardization of biological components and the design and construction of biological modules; (2) minimal genome research; and (3) design, synthesis, and assembly of the genome.
1. Standardization of biological modules and design and construction of biological components
The standardization of biological modules is to design and construct biological components (including promoters, terminators, protein encoding DNA sequences, etc.) according to certain standards or specifications, and to describe in detail about the biological components. When “parts” of synthetic biology—the parameters of biological components—are determined and standardized, the design and building of biological modules will become easy and reliable. The interaction relationship between inhibition and promoter of cell exhibits similar characteristics with switches and oscillators in the circuit system. Based on such characteristics, the design and assembly of the standardization biological element in different levels can generate similar systems of circuit. Such biological systems, the controllability and predictability of output can be improved to some extent. Biological module construction includes design and construction of new biological components, devices, or systems, and also includes redesign and transformation of existing biological systems in nature. Design or synthesis of some natural and nonnatural substances is also considered as one of the contents of synthetic biology, such as artificial synthesis of ribosomes.
2. Minimal genome research
A suitable carrier cell is required for the expression of new biological modules. Ideal carrier cells should have a streamlined genomic structure to reduce the complexity and improve the controllability and maneuverability of the designed system. The minimal genome is the minimum number of genes necessary for maintaining the growth and reproduction of the cells under the optimum conditions. Therefore, the core of minimal genome research is to determine the essential genes. According to the necessity of genetic information, we can purposefully streamline the existing genome and remove nonessential genomic fragments. In addition, we can also redesign, synthesize, and assemble essential genes. These two methods are now recognized as two strategies to achieve the smallest genome construction. Of course, this “minimum” is a relative concept. In addition, there is no universal minimal genome that can be used for various biological applications. With the development of DNA sequencing technology, more and more microbial genomes have been sequenced, which provides an important foundation for genomes minimization.
3. Design, synthesis, and assembly of genome
Synthetic biology research, whether construction of biological modules or minimal genome research, is the operation process of the genome sequencing. To achieve “engineered life,” the genome manipulation techniques, particularly genome sequencing technology and DNA synthesis technology, are essential. Genome sequencing technology allows us to “read” life’s “bible” and helps us understand the complex living systems. The establishment and improvement of the technical system is an important prerequisite for the development of synthetic biology.
In summary, the different synthetic biology researches interact with each other: the design and construction of biological components and standardization of biological modules allow us to realize purposeful design and transformation of life forms on the basis of an in-depth understanding of the complex life system; minimal functional genomics research provides the ideal expression vector for new designed biological modules; synthetic genome technology provides a solid technical support for previous two implementations. The mutual reinforcement of the three researches ultimately achieves new life forms of specific function and practical value.
Currently, industrial applications of synthetic biology have showed good prospects for development in the pharmaceutical, bioenergy, chemicals, and agriculture. The application of synthetic biology is an extension of the “cell factory” concept. Synthetic biology can address issues from energy, materials, environmental protection, and other social problems by the approach of artificial cell construction.
According to the synthetic biology principles, Zhang, Evans, and Mielenz (2007b) constructed a new catalytic system with 13 known enzymes, which use starch and water to produce hydrogen under mild conditions. The hydrogen is then used for electricity production by fuel cells, which realized the application of biohydrogen in the automotive fields. Keasling and coworkers transformed E. Coli by the methods of synthetic biology. The transformed E. Coli could produce complex biofuels (such as fatty esters, fatty alcohols, and waxes) by monosaccharide fermentation. Moreover, the transformed organism also had the ability to secrete hemicellulase to turn the hemicellulose into biofuels (Steen, Kang, & Bokinsky, 2010). Liao et al. reconstructed the isobutanol production pathway in E. coli and synthesize branched-chain higher alcohol biofuels using glucose (Atsumi, Hanai, & Liao, 2008).
In fact, the expression and regulation of genes and the complex metabolic networks inside the cells are as fine as spider webs and a small change will affect the whole body. The complexity of the expression and function of cellular gene and metabolism networks are much higher than the circuit board. Thus, even in today’s highly developed life sciences, the transplantation of streamlined minimal genome into mycoplasma without genetic materials is not successful. There is still a long way to go for synthetic biology.
5.3. Totipotent Cell Function: Artificial Cell
The biochemical conversion of biomass refining cell platform totipotent cell is to convert the substrate into a specific product with functional totipotent. The essence of the process is enzymatic catalysis in biological systems. Microbial cells have been rapidly applied to industry production for its diversity and simplicity. However, there are still some problems, for example: the permeability of the substrate membrane, which affects the final conversion rate; the side reactions leading to the degradation of substrate or products by passing reaction and the accumulation of byproducts. These problems, to some extent, limit the application of microbial whole cell transformation in industry.
From a molecular perspective, genes and proteins are essential to perform the original cell functions, and proteins are the carrier of cell function. Therefore, to build any type of cell, the proteins must be concentrated on. The enzymes are the executor of cell refining activities. Biomolecules, such as enzymes, are parceled or fixed in a semipermeable membrane that forms the artificial cells.
Enzymatic transformation and microbial transformation are important biological pathways in which energy, chemicals, and materials were produced with renewable biomass resources. Compared with the microbial processes, enzyme catalytic processes have advantages of clear transformation pathways, less by-products, simple operation, high yield, and easy process integration and optimization. By extracting enzymes involved in the desired product from microbial for the catalytic reaction, the production of byproducts can be limited, and the conversion efficiency of the objective product may be improved. Enzymatic processes have been widely used in industry, but there are still obstacles limiting broader enzyme use (Ma, Wang, & Su, 2009): (1) it is usually difficult to maintain the long-term stability of the enzymes in vivo because of the unsuitable extracellular environment, and it is easy to lose activity especially in the oil phase or oil–water interface; (2) the reaction rate is slower than that of chemical reaction; (3) since the enzyme requires extraction and purification and the repeat utilization is low, the cost for enzyme is high and even more so when the coenzymes are required in the system; and (4) the industrial applications of enzymatic process has been mostly confined to a single enzyme-catalyzed system, and multienzyme systems are still difficult, which limits the development of more high-value-added products.
In order to realize large-scale industrial applications of enzymes, the goal of efficient utilization of enzyme must be achieved. This requires not only the highest activity of a single enzyme play in each system (water phase, oil phase, the oil–water interface, etc.) but also the reuse of the enzyme and the coordination of multienzyme reactions and the coenzyme regeneration.
5.3.1. Multienzyme Reaction System
Most of the existing artificial biotransformations have been completed by a single enzyme and single cell, leading to the deficient efficiency. In nature, the biological energy conversion utilization is completed by multienzymes or cell systems, which is common in nature. Organism metabolism and biological symbiosis in nature are maintained by a variety of multienzyme systems. It is an efficient and accurate system, maintaining a high level of equilibrium between a series of enzymatic reactions. A microorganism multienzyme system in the catalytic process exhibits high selectivity, high efficiency, and a high degree of coordination, which provides a reference for establishing a multienzyme reaction system and optimizes the transformation of biocatalytic processes. The multienzyme reaction system has been used in a multistep completion of enzyme-catalyzed biochemical reactions in order to improve the overall yield of the reaction, shorten the reaction time, and reduce raw material and energy consumption.
At present, there are many reports about the use of multienzyme systems in biological synthesis of products, such as amino acids, organic acids, optically active alcohol ketones, nucleic acids, steroids, covering the fields of medicine, food, environmental protection. However, in domestic situations, it is only applied in starch, protein hydrolysis, and ethanol fermentation. Therefore, the multienzyme coupling reaction system has good development potential.
In biotransformation, according to the different sources of the enzyme, the multienzyme system can be divided into two types: the self-coupling reaction system and the interspecies coupling reaction system (Xie, Zhou, & Gao, 2004).
5.3.1.1. A Self-coupling Reaction System
The self-coupling reaction system refers to achieving the coupling of an enzyme reaction with the cooperation of multiple enzymes in microorganism. This system is to obtain a single strain which has high multiple enzyme activities by screening manually, or to obtain the high activity strain by recombinant expression of different enzyme genes derived from different source in the same strain, so as to achieve the coupling of the intracellular response and the efficient conversion of the target product.
Only one kind of microorganism was used in the self-coupling reaction system with the demand of a variety of enzymes with high activity in the microorganism. Moreover, the overall increase of the activity of multiple enzymes is necessary for improvement of the efficiency of the entire system. Although the coexpression of different enzymes can be achieved in the same strain by gene recombination technology, it is quite hard to transform due to the conservative nature of the genetic mechanisms of microbial cells. Thus the application of the self-coupling multienzyme reaction system has been limited.
5.3.1.2. Interspecies Coupling Reaction System
The interspecies coupling reaction system refers to the fact that the enzymes required for the reaction exist in various microorganisms, which constructs coupling reactions among heterogeneous microorganisms. The interspecific coupling reaction systems can be divided into two kinds: cofactor regeneration coupling system and substrate coupling reaction system. The cofactor regeneration coupling reaction is only completed with the participation of cofactor ATP and NAD(P)H and performed through the coupling reaction of cofactor regeneration. In the substrate coupling multienzyme reaction system, it could be started from the compounds with extensive sources and low cost, and a synthetic intermediate is obtained by enzyme catalysis. Then the intermediates are catalyzed by another enzyme to obtain the desired product. The system can reduce the separation and extraction steps of intermediates, shorten the reaction process, and then greatly reduce the separation costs and environmental pollution.
The interspecies coupling reaction system compared with the self-coupling multienzyme reaction system has the following advantages. First, nicotinamide is the key coenzyme of electron transfer process in biological metabolism in the construction of the cofactor regeneration system and the metabolism and reproduction is precisely regulated by the microorganism itself. In a single cell, the nicotinamide acts on each reactions with a fixed ratio and it is hard to ensure recycling by using single cell self-coupling multienzyme systems. As for the interspecies coupling multienzyme reaction system, there is no correlation between growth and metabolism of different microorganisms. It is possible to make a successful coupling reaction among the multienzymes by regulation. The successful application of nicotinamide cofactor regeneration system realizes the large-scale preparation of chiral alcohol. Second, the enzyme required for the reactions are from different microorganisms, the choice of microorganisms required for building multienzyme coupling system has great diversity and flexibility. Meanwhile, the interspecies coupling reaction system can increase the activity of specific enzyme of different microorganisms by gene recombination technology and lower technical difficulty. The current research about multienzyme coupling reaction systems focuses on interspecies coupling reaction system for its better development potential.
Learning from a highly coordinated multienzyme reaction system of microorganisms in nature, the multienzyme system will tend to a highly coordination of various enzymatic reactions by using genetic engineering technology and directed enzyme evolution technology, which will also promote the industrial process of the multienzyme system’s application in amino acids, organic acids, and pharmaceutical intermediates and other fine chemicals.
5.3.2. Carrier Immobilization
Cell immobilization is developed on the basis of immobilized enzyme technology. The free cells are fixed on the defined spatial region (a suitable insoluble carrier) by physical or chemical methods, remaining activity of enzymes and can be used repeatedly.
In 1959, Hattori and Furusaka used resin to adsorb E. Coli to realize the immobilization of cells for the first time, which showed broad development prospects in the fields of energy, environment, food, medicine, and chemistry. Cell immobilization is gradually applied in biochemical biomass conversion. Shen et al. coimmobilized Aspergillus niger spores with rich cellobiase and Lactobacillus delbrueckii in calcium alginate gel beads, coupling fixed-cell systems and enzymatic hydrolysis system of cellulosic feedstock. With this new bioreactor for lactic acid fermentation, the results of repeated batch reactions assay showed that synergistic coimmobilized cells have sustained, stable, and efficient lactic acid production capacityand can be used repeatedly (Shen & Xia, 2008).
5.3.2.1. Conventional Immobilization Methods
Currently microbial immobilization methods are mainly adsorption, embedding, cross-linking, and covalent binding methods (Wang, Huang, & Luo, 2007).
The adsorption method uses charge between the electrostatic microbial cells and carriers to immobilize microorganism cells. The method is simple; reaction conditions are mild; the process of fixing microbial cells has little influence on its activity. However, the combination is not firm, cell is easy to shed, and the number of fixed cells is limited by the type of carrier and its surface area.
The embedding method is to embed the microbial cells in a water-insoluble polymer gel pore network space formed by polymerization, precipitation, or changing the solvent, temperature, or pH. A gel polymer network can prevent the leakage of the cell, allowing diffusion of the substrate and the product. This immobilization method has the advantages of easy operation, is simple to operate, fine multienzyme systems maintaining performance, and less impact on the activity of microbial cells, which makes it one of the most widely studied methods for immobilized microorganism.
In the cross-linking immobilization method, the microorganism cells react with two or more polyfunctional reagent to form a covalent bond. The operation conditions are intense and have a great influence on the activity of microbial cells.
In the covalent bonding methods, the functional groups (such as amino, carboxyl, mercapto, hydroxy, imidazolyl, and phenolic groups, etc.) on the surface of microorganisms form chemical bonds with carrier chemical groups. The linkages between microbial cells and the carrier are very strong; however, the reaction conditions are intense and hard to control.
5.3.2.2. Choice of Carrier
There are multiple characters that make ideal microbial immobilization carriers. First, long life, high mechanical strength, high capacity, low price. Second, not easy to be biodegraded; simple immobilization process, easy to shape at room temperature, nontoxic to microorganisms during immobilization process and after immobilization. Third, good biochemical and thermodynamic stability; good matrix permeability; good precipitation separation; no interference in the function of biological molecules. The key of cell immobilization technology is the performance of the immobilization carrier material. Immobilization carrier materials currently used are mainly organic polymer carriers, inorganic carriers, and composite carriers (Qu & Yue, 2007).
5.3.2.2.1. Organic Polymer Carrier
Organic polymer carriers include natural polymer carriers and synthetic organic polymer carriers.
1. Natural polymer carriers are generally nontoxic to microorganisms, have good mass transfer performance, but low strength and can easily be broken down by microorganisms in anaerobic conditions. Common natural polymer carriers are agar, gelatin, carrageenan, sponges, chitin, alginate, and chitosan. The agar-embeding cell method is simple and nontoxic to cells, with a larger clearance, allowing the diffusion of high molecular substances, but the mechanical strength and chemical stability is not good.
Alginate is a widely used immobilization carrier. It has good chemical stability, nontoxic, high efficiency, and is suitable for fixing living cells or sensitive cells, but it cannot resist phosphate and Na+, K+, Mg2+, and other cations with high concentrations, and is broken easily and dissolved. The gel strength is not enough and it is hard to repeat the use of immobilized cells.
2. Synthetic organic polymer gel carriers show good resistance to antimicrobial decomposition, high mechanical strength, and good chemical stability. However, the mass transfer performance is poor and the cell activity will reduce in the embedding process. Common synthetic organic polymer gel carriers are polyacrylamides, light-curing resins, polyvinyl alcohol, and polyacrylic acid gel.
Polyacrylamide (ACAM) gel has good mechanical, chemical, and thermal stability. Due to the toxicity of acrylamide monomer to cells and heat release of cross-linking process, cells often inactivated during immobilization process. Thus, there are few studies about the use of ACAM as embedding agents. However, the cells can overcome this disadvantage by first being embedded with agar and then ACAM, which is called secondary fixation.
Polyvinyl alcohol (PVA) has high gel strength, good chemical stability, and strong antimicrobial decomposition performance. Compared with the ACAM gel, its biological toxicity is very small and causes little harm to the activity of cells. It is the most widely studied immobilization carrier.
5.3.2.2.2. Inorganic Carrier and Composite Carrier
The inorganic carriers, such as porous ceramic beads, crushed red brick, sand, porous glass, kaolin, diatomaceous earth, activated carbon, and alumina, usually have a porous structure, and fix microorganisms or cells with effect of adsorption and charge. Those carriers have advantages of high mechanical strength, nontoxicity to cells, and high resistance to biodegradation. Composite carriers are the combination of organic carrier materials and inorganic carrier materials, and have the advantage of performance complementarity.
5.3.2.2.3. Microcapsule Immobilization
Among most immobilization systems, microcapsule immobilization has taken much attention (Ma, Lin, & Yao, 2010). A suitable liquid environment is wrapped by a layer of microcapsule membrane. Various cells grow coordinately in the internal to achieve a variety of biochemical reactions. The outer membrane has the functions of isolation, protection, and mass transfer. Thus, the microcapsule system is like a virtual “cell factory,” absorbing nutrients from the outside, synthesizing specific products by the internal complex metabolic reactions, and finally discharging the products outside the membrane.
The cellulose sulfate sodium/ploy-dimethyl-dially-ammonium-chloride microcapsule system has good biocompatibility, simple preparation, good physical and chemical properties stability, and good mechanical strength. By using this system to study immobilization and culture of varied cells with the products including ethanol, lactic acid, glutamic acid, 1,3-propanediol, and thrombolytic enzymes, it was found that the system was especially suitable for anaerobic culture.
5.4. Intercellular Synergy
Microbes are the core of industrial biotechnology. Fermentation requires microorganism strains with good performance. Early research on microbial fermentation transformation was screening of natural high-yield strains, followed by obtaining a mutant strain with increased fermentation capacity by chemical mutagenesis and high-throughput screening technologies. The great limitations of these traditional strain breeding technologies was that the microbial fermentation products of mutant species are very limited, which are ethanol, acetone, butanol, glycerol, organic acids, amino acids and antibiotics, and other metabolites. Therefore, researchers began to pay attention to the mixed culture of transformation process.
In the platform of a refining cell, two or more microbial cells play a role of synchronization for competitive advantage, and the effect is much better than that of individual microorganism cells. The specific phenomenon is called synergistic effect between the microbial cells, which is the mixed culture of microorganisms. In long-term experiments and production practice, people gradually find that many important biochemical processes must be completed by two or more cocultured microbes.
Microbial fermentation is widely used in the production practice, which can replace the production of single fermentation in many cases. Some new products are possibly produced during the mixed culture process because the mixed microbial fermentation is a biological hybrid system and the microorganisms in the system have a coordinating role on the growth and metabolism. Single strain fermentation has low utilization efficiency of raw materials, equipment, and energy while multistrain mixed fermentation can overcome these shortcomings. However, study on mixed fermentation and optimization of culture conditions is limited.
5.4.1. Characteristics of Mixed Fermentation
1. To obtain certain products those are not available for single fermentation
Pure fermentation accounts for a large proportion of the modern fermentation industry, but in long-term production practice, it is found that some products can only be the product of multistrains mixed fermentation. The two-step fermentation process has been widely used in the production of vitamin C and the second step, from l-sorbose to keto-l-gulonic acid, is a mixed fermentation process of two bacteria. One bacterium is Gluconobacter oxydans and the other bacterium is Bacillus megaterium. The traits of the two bacteria are different, and their role is also not the same. Either bacteria cultured alone is not enough to produce cologne acids, or only two bacteria mixed fermentation can successfully complete the conversion process of cologne acids.
2. To promote the product yield
Mixed fermentation uses symbiotic effects or nutritional interactions between two or more bacteria, and overcomes the cumulative adverse effects caused by accumulation of intermediate. After combinations of different strains, the activity of enzymes is greatly increased. This is because the mixed fermentation takes the advantage of effects from different strains, improves the enzyme production capacity, and increases the yield of products.
Cellulase production by mixed fermentation proves to be an effective way to utilize natural cellulose resources. Fungi are used as cellulase production strains in industry and the produced cellulase is generally secreted into the medium. It could be easy to obtain cell-free enzyme products by filtration and centrifugation. Taking cellulose production by fungal mixed fermentation as an example, the mechanism of mixed fermentation is summarized (Zheng, 2011) as follows:
a. Weakening feedback inhibition of enzymes by mixed fermentation
During the enzymatic hydrolysis of cellulose for the production of glucose, glucose and cellobiose have strong feedback inhibition on cellulase, which affects the rate and extent of hydrolysis of cellulose. If the bacteria that can break down the cellulose and the bacteria which can utilize glucose and cellobiose are mixed together during one fermentation process, the feedback inhibition from glucose and cellobiose is greatly diminished. So, for the decomposition of cellulose, mixed fermentation is faster and more thorough than single pure culture fermentation. Si-Mei et al. studied the effects of Candida sp. on the activity of cellulase and amylase during solid-state fermentation of Aspergillus niger and Aaspergillus fumigatus. The results showed that the inoculation of small amounts of Candida sp. can greatly improve the activity of cellulase and amylase because yeast use cellobiose and other small molecule sugars formed by hydrolysis, reducing the inhibition of cellulase and amylase synthesis by cellobiose, and thereby increasing the activity of fermentation products (Si-Mei, Xue, & Cai, 2002).
b. Enhancing total enzyme activity by enzymes complementary
In the study of fungal cellulase fermentation process, it is found that although Trichoderma sp. is recognized as the best cellulase-producing bacteria, there are still disadvantages in two aspects, one of which is the suspected toxicity and the other is the low activity of β-glucosidase. Many Aspergillus sp., such as Aspergillus niger, can produce β-glucosidase with high activity and are recognized as strong and safe strains for cellulase production. The studies of mixed culture of Trichoderma sp. And Aspergillus sp. indicate not only that β-glucosidase activity increased but also that the endo-cellulase and cellobiohydrolase activity are improved.
c. Mutualism
Cellulase-producing strains mostly include Trichoderma koningii, Trichoderma viride, Aspergillus fumigatus, and Aspergillus niger; lignin-decomposing bacteria include Sporotrichum sp. and white-rot bacteria; protein enhancing bacteria are usually yeast, such as Candida utilis, Candida tropicalis, and S. cerevisiae. The mutualism of strains puts the enzymes in a coordinating ratio and greatly improves their activity. Pu et al. cocultured nitrogen-fixing bacteria (Azotobacter sp.) and cellulose decomposing bacteria (Trichoderma sp.) and observed that the growth of the two strains and nitrogen fixation were higher than these of a single culture. The reason is that nitrogen-fixing bacteria can not only fix atmospheric nitrogen but also produce different vitamins and auxin, which can stimulate crop growth and development and strengthen other rhizosphere microbial life activity (Pu, Zhong, & Zheng, 2000). About half of the organic matter in soil is cellulose; cellulose-decomposing bacteria can break down these substances into simpler substances which can be used as a carbon source by nitrogen-fixing bacteria.
d. Increasing yield
Tu et al. studied the effects of two mixed Aspergillus strains on the activity of three cellulase components. The results showed that the two fungi were inoculated by a certain percentage, the activity of three cellulase components would greatly improve than single fermentation. The activity of FPase, microcrystalline cellulase, and carboxymethyl cellulase increased by 2.2∼51.1%, 20.7∼332.6%, and 29.4∼29.6%, respectively (Tu, Xue, & Si-Mei, 2004).
5.4.2. Precautions of Mixed Fermentation
Compared with single microorganism fermentation, mixed fermentation can take advantage of microbial diversity to achieve synergy among bacteria and improve the conversion rate and yield as much as possible. However, the following details must be carefully considered (Song, Zhou, & Chen, 2008; Zhang & Hou, 2010).
5.4.2.1. Strain Combination
The combination of mixed fermentation bacteria usually includes cellulose-decomposing bacteria, lignin-decomposing bacteria, and protein-enhancing bacteria. Cellulase-producing strains mostly include Trichoderma koningii, Trichoderma viride, Aspergillus fumigatus, and Aspergillus niger; lignin-decomposing bacteria include Sporotrichum sp. and white-rot bacteria; protein-enhancing bacteria are usually yeast, such as Candida utilis, Candida tropicalis, and S. cerevisiae. It is necessary to pay attention to the compatibility of different strains and try to take advantages of microbial strains with similar habits during strain combination.
5.4.2.2. Different Fermentation Conditions
Fermentation temperature, pH, time, moisture, and other factors and their interactions have a significant influence on fermentation. Temperature is the primary factor for solid-state fermentation. The starting medium pH and moisture content must first be adjusted according to the characteristics of the strains used and raw materials; then the temperature suitable for the growth and reproduction of microorganisms should be set; finally the best culture time for higher yield of aim products should be determined.
5.4.2.3. Pretreatment of Fermentation Feedstock
Since the feedstock for general mixed fermentation is lignocellulosic biomass resources, the pretreatment of feedstock is necessary to separate cellulose, hemicellulose, and lignin for efficient enzymatic reactions. The pretreatments usually used include cooking, acid/alkali treatment, organic solvent-treatment, steam explosion, and the wet oxidation methods. In addition, the nutritional content of the lignocellulosic feedstock does not fully meet the needs of microbial growth and reproduction, so we have to add the appropriate carbon source (such as bran) and nitrogen (such as urea) in order to increase enzyme activity and protein content of the product. From the point of the industrialization, the conditions for mixed fermentation are simpler and easier to operate compared with pure strain fermentation. However, the mixed fermentation system is more complex than pure strain fermentation and the existing studies should be more thorough and extensive.
5.5. Construction of a Cell Refining Factory
Cell refining makes use of complex microbial metabolic networks and regulatory networks, decomposes biomass, and synthesizes a range of energy products to replace fossil and chemical products. A microbial cell factory is the core technique of cell refining.
The so-called “factory” is the place that can produce or manufacture a product. Therefore, the “factory” in a general sense should have specific production lines, and the corresponding power and other auxiliary systems, and is normally operated in certain management programs. The elements of a “factory” are designed based on what people want. The production line and auxiliary system are designed based on needs and regulated by the production schedule. “Cell factories” also have corresponding components (Zhang, Li, & Ma, 2007a). The so-called cell plants aim to discover the nature of genes, proteins, and metabolic networks, and reallocate material flow and energy flow systems of microbial cell metabolism by artificial restructuring and optimization.
Microbes are the most widely distributed species on earth, and their metabolic diversity is determined by species diversity and genetic diversity. Microbes play a critical role in the earth’s irreplaceable material cycle and have formed varied mechanisms for nutrition taking in long-term evolution. Microorganisms degrade the biomass by secreted enzymes and convert biomass to bioenergy (biohydrogen, bioethanol, biodiesel, etc.), important chemical raw materials (such as ethylene, 1,3-propylene glycol, etc.), polymers, and other products. Of course, the compositions of biomass materials are abundant, including a variety of substances with hydroxyl, carbonyl, benzene, and other oxygen-containing groups. Compared with petroleum feedstock containing —(CH2)n— linear polymeric structure, biomass can provide more opportunity for development of new products, which are more conducive to the produce of various chemicals by chemical transformation (Chen & Wang, 2008), shown in Fig. 5.5:

Figure 5.5 Products from cell refining factory.
The production capacity of a biological cell factory is the most important factor to determine its ability to compete with petrochemical manufacturing. An efficient cell factory must have the following six important characteristics:
1. The cell should have a clear physiological genetic background and good genetic stability, as well as being easy to genetically transformation. Therefore, the construction of cell plants is mostly based on model microorganisms (such as E. coli) whose genetic background is relatively clear.
2. Fast growth, strong anabolic ability, high production yield, high production rate, and yield. Thus, it is possible to reduce the costs of biological manufacture.
3. Simple fermentation process, reducing the equipment cost, and operating costs.
4. Utilization of simple inorganic salt medium and inexpensive carbon sources to thereby reduce production costs.
5. Using cellulose as raw material.
6. Superior physiological performance, tolerance to high temperature, high osmotic pressure, high product concentration, and low pH.
5.5.1. Modification of Microbial Cell Properties
The biorefining is parallel to petroleum refining, converting biomass to fuel, materials, or compounds platforms other types of chemical with the help of the natural ability of the microorganism and recombinant microorganism cells and a series of biochemical pathways (similar to unit operation, such as petroleum refinery, cracking, hydrocracking, reforming). It needs to modify the characteristics of microbial cell to meet industrial production needs.
5.5.1.1. Optimization of Strain Tolerance and Reduction of Metabolic By-products Synthesis
The microbial fermentation production process inevitably suffers some inhibition of adverse environmental factors on product synthesis. The resistance of cells to an adverse environment is a very complex phenotype. By transcriptome comparison of strains grown in different environments, genes that are closely related to the phenotype could be found, allowing researchers to better enhance the strain tolerance to adverse environmental by metabolic engineering. Hirasawa et al. (Hirasawa, Yoshikawa, & Nakakura, 2007) compared the transcriptome differences in two strains of S. cerevisiae which had different resistances to ethanol. By using cluster analysis, they found that the expression level of tryptophan biosynthesis genes has a close relationship with ethanol tolerance. Overexpression of tryptophan biosynthesis genes could make strains with low tolerance to ethanol have 5% (V/V) ethanol tolerance ability and exogenous tryptophan addition also enhanced expression of tryptophan permease and increased ethanol tolerance. Hirasawa et al. (Hirasawa, Nakakura, & Yoshikawa, 2006) also analyzed transcription of two S. cerevisiae strains with different osmotic tolerance under high salt conditions and found that the sodium ion pump and copper metallothionein gene were closely related to cell osmotic resistance. Increasing the expression level of these genes can significantly improve osmotic resistance of strains. Glycerin is a by-product when S. cerevisiae was employed for ethanol production by anaerobic fermentation. Under anaerobic conditions, the formation of glycerin converted NADH into NAD+. The glycerol-3-phosphate dehydrogenase encoded by the gene GDP2 and GDP1 converted dihydroxyacetone phosphate into glycerol-3-phosphate. The breaking of GDP2 gene reduced the production of glycerol but slowed down the cell growth rate. In addition, strains with GDP1 and GDP2 gene double deletion cannot grow under anaerobic conditions. Thus, Nissen et al. constructed a new way to produce NAD+ in Azotobactervinelandii with GDP1 and GDP2 double gene deletion, thereby reducing the formation of glycerol by 40% (Nissen, Kielland-Brandt, & Nielsen, 2000).
5.5.1.2. Expanding Range of Substrate Utilization
Expanding the range of substrates utilization has significant importance for the production of biobased chemicals by biomass, phenotype of fast utilization (especially under anaerobic conditions) of xylose, galactose, and other substrates in some microorganisms are related to the complex changes in gene expression. Bro, Knudsen, and Regenberg (2005) analyzed S. cerevisiae strains with different galactose uptake rate by transcriptome and identified PGM2 genes encoding glucose phosphate mutase, which were taken as new targets. By enhancing the expression of the gene, the galactose uptake rate of engineered bacteria was increased by 70%. The study also showed that transcriptome analysis of strains with different substrate consumption rates would greatly improve the efficiency of obtaining useful information. Bengtsson, Jeppsson, and Sonderegger (2008) analyzed the transcription differences between the normal group and four S. cerevisiae strains with different xylose-utilizing ability, and found that the expression of 13 genes in the four strains had changed. In the normal strains with corresponding overexpression or deletion of these different genes, it was found that there were five genes that could effectively improve the xylose utilization capacity of the strain.
5.5.1.3. Production of Heterologous Metabolites
Eliasson, Christensson, and Wahlbom (2000) separated XYL1 and XYL2 genes from P. stipitis. Encoding of the two genes was dependent on NAD(P)H-dependent xylose reductase and NADH-dependent xylitol dehydrogenase. After being inserted into the S. cerevisiae, the endogenous XKS1 gene (encoding the xylulose kinase) was overexpressed, which allowed the S. cerevisiae strain TMB3001, growing under anaerobic fermentation and producing ethanol. Meanwhile, the overexpression resulted in a reduction of xylose utilization and the ATP/ADP ratio (Toivari, Aristidou, & Ruohonen, 2001).
5.5.2. Design of Cell Refining Factory
In order to obtain efficient microbial cell factories, the three steps of design, build, and optimization are needed. After clarifying the product goals, the first question is how to design a highly efficient cell factory to produce the desired product. Most microbial fermentation products are a mixture of compounds, so it needs to be reformed on the basis of systematic research on microbial metabolic networks. The most effective way of systematic research on microbial metabolic networks is to construct metabolic network models. Determination of microbial genome sequences and the development of a gene function annotation tool allow us to construct a genomic-level metabolic network model, which greatly enhances our ability to analyze the metabolic network structure. So far, scientists have completed approximately 20 microbial metabolic network models. These models enable us to recognize the complexity of microbial metabolic networks in the systemic level, to predict the physiological properties of the cells and the genetic changes or the metabolic response of cells after environmental disturbances, and to simulate the genetically engineered target genes, and provide a good foundation for the cell plant design.
5.5.3. Construction of Cell Refining
Establishment of the metabolic network model and computer simulation analysis provides the basis for the design of the cells’ factory. The specific implementations of cells construction will be completed by genetic modification. Common means of genetic manipulations include gene knockout, gene effective expression, gene integration, gene expression regulation (transcriptional level and posttranscriptional level); gene knockout and integration are two powerful tools widely used in building microbial cell factories. Knockout can be used in inactivation competition pathway, so that more metabolic flux flows into the target products. Integration of the gene can introduce a fragment of an exogenous gene into the cells’ factory; the foreign gene can assign the cells a new capacity, synthesis of new product, and enhance the resistance of cells to enable the production of products which could not be previously synthesized. Regulation of gene expression is another common genetic transformation strategy. By constructing a library of promoters with different expression levels, and controlling the posttranscriptional process, such as termination of transcription, mRNA degradation, and translation initiation, the intensity of gene expression can be regulated to control the distribution of metabolic flux and then more effectively to synthesize the target products.
Li et al. achieved the conversion of nonmethylated plasmid in Clostridium acetobutylicum by inactivation of restriction modification systems. They further analyzed the existing deficiencies of the homologous recombination based on nonreplicating plasmid, homologous recombination based on replicating plasmid, antisense RNA technology, and type II intron gene inactivation technologies, and pointed out that improving the conversion efficiency is the key breakthrough of the genetically engineered operating system of Clostridium acetobutylicum. Finally, they provided suggestions about how to develop an efficient genetic manipulation system of C. acetobutylicum (Jia, Zhu, & Zhang, 2011).
The microbial DNA transformation method is to import foreign genes and the current microbial transformations are mainly chemical and electrical transformation methods. These methods have shortcomings, such as the long time required for preparation of competent cells, cell activity loss during treatment, and incubation after treatment. Tan et al. (Tan, Wang, & Zhao, 2010) developed a sepiolite-based microbial DNA transformation method. Sepiolite is a mineral nanomaterial, which is cheap, abundant, and harmless. This transformation method does not require the preparation of competent and incubation after treatment, to obtain a higher conversion rate than the conversion of calcium transformation process.
The genome mixed group is a new method for cell factories transformation. Genomes from different strain sources can be fully recombined by means of protoplast fusion cycle, which increases the opportunity to integrate the mutation into recombinants. Yang, Fan, and Xiao (2010) investigated the effect of genome mixed group changes during multiple rounds fusion process of Bacillus subtilis. By comparing the mixed result of Streptomyces coelicolor and Lactobacillus sp. genome groups, and combination with computer simulation of cycle fusion process, they demonstrated that the high frequency recombination technique of microbial cells is the basis on which to improve more sufficient genome mixed effects of Bacillus subtilis.
Surface display is a valuable genetic manipulation technique, which allows the expression of exogenous peptides to show on the cell surface in the form of fusion protein. Spore surface display technology as a microbial surface display technology has attracted much attention from researchers because of its unique advantages, such as heterologous proteins expressed without transmembrane process and the resistance of spore. Xu, Wang, and Ma (2010) described the physiological structure and the formation of spores, construction principles of spore surface display system, and types of spore surface display system currently constructed. Bacillus subtilis surface display technology will get more attention in the field of biocatalysis and cell factories due to its safety feature. In addition, Bacillus subtilis also provides an important reference for the industrial production of high-value-added compounds.
5.5.4. Optimization of Cell Refining Factory
By designing and constructing the plant factories, the primary cells’ factory can be obtained. However, its production capacity and physical properties have many shortcomings. A cells’ factory is an artificially designed microbial metabolism system. The introduction of multicell genes or an entire metabolic pathway may lead to some problems, such as metabolic imbalance, accumulation of intermediate metabolic products, and cannot achieve the requirements of industrial production. Thus, it is necessary to optimize the cell plants. Jiang, Li, and Ma (2010) described the coordination and optimization of multigene expression strategies from levels of transcription and translation. Components optimization by adding artificial scaffold protein is a new optimization technique of cell plants that has been recently developed. The technique can precisely control and optimize stoichiometric ratio of multiple enzymes in the metabolic pathway, balance the flow of each part in the pathway, shorten the space between each enzyme, and form the substrate channel, thereby greatly enhancing the efficiency of biosynthesis in the cell plant.
Microbial synthesis of high-value-added fine chemicals is a relatively mature technology. These processes usually have fine produce condition and are in a small-scale production. Therefore, biosynthesis has certain advantages. The biorefinery process for bulk chemicals production is featured by large amounts of output and simple medium composition and operation process. These requirements reduce the nutritional needs of the industrial strains and improve their tolerance to substrates and products, enhancing industrial adaptability of cell factories. In recent years, the researches of extreme environmental microbes have laid a foundation for the industrial adaptability transformation of cell factories. However, to fully meet industry needs and realize industrial applications is still an important direction for future research on plant cells. In order to realize the applications of cell plants in the field of biorefinery, further studying and improving industrial adaptability of cell plants are always necessary, which could really develop cell plant manufacture into a high-yield, efficient, and economical modern industrial biotechnology industry.
Among them, the further industrial development of biochemical transformation is inseparable from the core production unit-bioreactors. The reactions in the bioreactor occur at multiple scales, such as the genetic characteristics at a molecular level, the regulation of cell metabolism at a cell level, and mixing and transfer at a reactor level. Therefore, how to use the multiparameter detection technology of bioreactor and online computer control and data processing technology to associate phenotypic data with genetic structure relate to metabolic regulation, is not only important content of process optimizing and amplification of reactors but also the original intellectual property technologies.
In summary, the development of efficient microbial cell plants can greatly enhance the production capacity and physiological performance of current industrial microorganisms, and reduce production costs and expand the products varieties of biomanufacturing. With the rapid development of new technologies, microbial cell plant techniques will also be more perfect and greatly promote the industrial upgrading of traditional petrochemical manufacturing industries, making tremendous contributions to the sustainable development of human society.
..................Content has been hidden....................
You can't read the all page of ebook, please click here login for view all page.