
Chapter 4
Enzymatic Hydrolysis of Pretreated Biomass
Abstract
The key technologies in biochemical biomass conversion are how to degrade complex polysaccharides in the biomass polymer into monosaccharides efficiently and convert monosaccharides with special functions into biobased products. Compared with physical–chemical treatment methods, enzymatic hydrolysis method holds many advantages, such as mild conditions, no toxic degradation products, high sugar yield, low equipment cost, and high specificity. It plays an irreplaceable role in biological conversion of biomass. The previous section has carried out a detailed discussion on the physical structure and chemical characteristics of biomass resources. It lays the foundation for further enzyme conversion and how to choose an effective pretreatment method to eliminate the degradation barrier, and expose enzyme loci. The enzymes adopted/applied in the biomass conversion will be discussed in detail in this chapter. Furthermore, it will also be illustrated how to build and exploit the enzyme platform to realize the efficient biomass conversion.
Keywords
enzymes platform
physical structure
chemical characteristics
4.1. Reviews on the Enzymes Participating in the Biomass Degradation Process
The essential enzymes for the biomass degradation process mainly include cellulase, hemicellulase, the lignin degradation enzyme system, and cutin enzyme and cellulose enzyme cofactors. Establishment of the biomass biochemical transformation platform mainly refers to the preparation and application of various enzymes, especially application of the synergy effect between different enzymes. The biomass degradation process requires enzymes cofactor, cutin enzyme, and cellulose enzyme solution. The biomass biochemical invertase platform mainly refers to the preparation and application of various enzymes, especially the synergy effect among different enzymes.
4.1.1. Cellulase
Enzymes with cellulose degradation activity are mainly produced by aerobic fungi and anaerobic bacteria. In the tropics, termites and some insects can also degrade cellulose, and most of them coexist with the cellulolytic microbe. Cattle, sheep, deer, and other ruminants are also important cellulose decomposers, in whose rumens cellulose gets degraded by coexisting bacteria (Jing, 2010).
Because of the strong interactions, such as hydrogen bonding and hydrophobic accumulation between adjacent molecular chains in the natural cellulose crystal area, natural cellulose is difficult to degrade. Therefore, the specific activity of single cellulose enzyme is much lower than others. When acting on insoluble substrates, the reaction rate is no more linearly related to time and enzyme, the reason for which remains unknown. Early researches held views that the cellulose could only destroy cellulose crystalline structure and C1enzymes with hydrolysis activity and Cx enzymes for β-1,4 glycosidic bond decomposition did not exist. After the 1970s, due to the development of biochemistry and molecular biology, the fungal cellulose enzyme system represented by Trichoderma reesei was elucidated gradually (ZhenJiang, 2007).
Fungal cellulases generally contain the following three components: (1) 1,4-β-d glucan hydrolase. These enzymes can randomly degrade the β-1,4 glycosidic bond within the cellulose molecules; (2) 1,4-β-d-glucan cellobiohydrolase enzymes (1,4-β-d-Glucan cellobiohydrolases, CBHs, EC 3.2.1.91,) referred to the cellulose exonuclease (exoglucanases), which can cut the glycoside keys at the reducing or nonreducing end of the molecule and generate cellobiose; (3) β-glucosidase (β-d-glucosideglucohydrolases, β-glucosidases, EC 3.2.1.21, referred to as BG), these enzymes degrade the cellobiose into individual glucose molecules (Gao, 2003).
There are three hypotheses on the mechanism of each cellulase component: the C1–Cx hypothesis, sequential action hypothesis, and synergy model (Gao, 2003; Lian & Wang, 2007).
Reese proposed the C1–Cx hypothesis to explain the action mode of cellulase enzymes in 1950 (Reese, Siu, & Levinson, 1950). The hypothesis holds that firstly, the C1 enzyme acts the crystalline of the cellulose and makes it adaptable for the C2 enzyme. The C1enzyme can randomly hydrolyze crystalline cellulose, glucose, soluble cellulose derivatives, and β-1,4-oligomer; β-glucosidase converts cellobiose, and cellotriose sugar into glucose. In the hydrolysis of natural cellulose (high crystallinity cellulose), the C1 and Cx enzymes coordinate to act in different stages: the first stage is that C1 enzyme acts on the crystalline cellulose, but there is only conversion, not hydrolysis. Then Cx enzyme hydrolysizes converted amorphous cellulose partly into soluble monosaccharides.
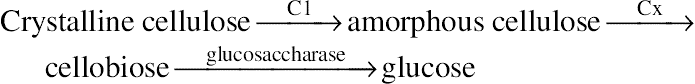
However, Reese et al. proposed that C1 and Cx hydrolyze cellulose in different phases, that is, it is only when C1, Cx, and β-glucosidase coexist that natural cellulose can be hydrolyzed. Yet, it has not been verified by the experiments, since if we firstly let C1 enzyme act on the substrate and then separated C1 and added Cx, the crystalline cellulose could not be hydrolyzed.
Sequential action hypothesis on the mechanism of action of cellulase holds that exoglucanase (CBHI and CBHII) firstly hydrolyzes insoluble cellulose fiber into soluble dextrin and cellobiose and then the endoglucanase (EGI and EGII) acts on the fiber dextrin to generate cellobiose. Finally, the BG decomposes the cellobiose into two glucoses, which is shown in Fig. 4.1.
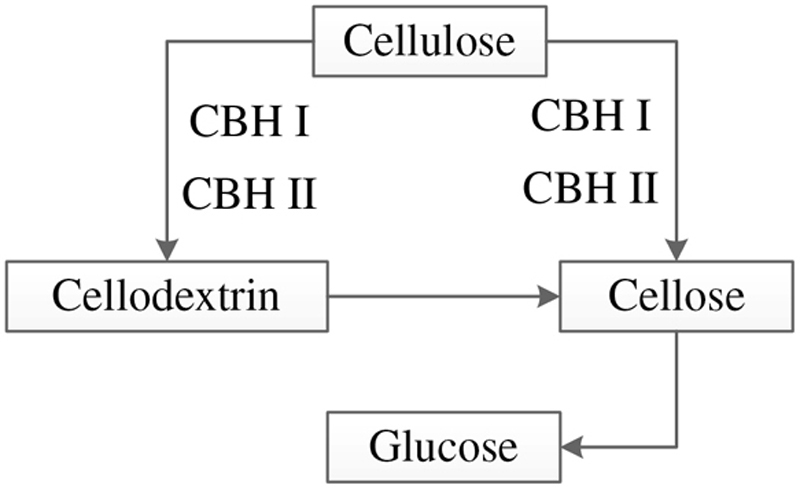
Currently, the synergy model is generally accepted for the explanation of degradation mechanism of cellulose, which is shown in Fig. 4.2 (Woodward, 1991).

Figure 4.2 The synergistic degradation model.
4.1.2. Hemicellulase
Hemicellulase usually refers to the enzyme mixtures acting on noncellulose and nonpectin biomass. They can be divided into two categories: depolymerase and debranching enzymes. Depolymerase acts on sugar chain backbones and also can be divided into two categories: enzyme I have the endo activity acting in the middle of polymers; enzymes II have the exoactivity acting from the end of polymers. In fact, many enzymes own two kinds of activities at the same time. In addition, a series of oligosaccharide-producing enzyme systems can be achieved by combination of endo- and exoenzymes, such as β-glucosaccharase (3.2.1.21), β-xylan glucosidase (3.2.1.37), and β-mannosidase (3.2.1.25). Due to its low polymerization degree, it is difficult to distinguish between the endo- and exoaction mode.
The debranching enzyme is a kind of auxiliary enzyme, which can be divided into enzymes acting on ester bonds and glycoside bonds. The former ones include α-l-arabinofuranosidase (3.2.1.550) and α-glucuronidase (3.2.1.39); the latter include acetyl xylan esterase (3.2.1.72) and ferulic acid enzyme (3.2.1.73), which mainly act on xylan. It is reported that hemicellulase enzymes also contain the enzymes acting on acetylated polysaccharide glucomannan, including glucomannan and galactomannan.
Only part of hemicellulose enzymes exhibit cross activities toward different hemicellulose, most of which possess strict specificity, and only acting on particular sequence of oligosaccharides. For example, β-xylosidase is suitable for the hydrolysis of xylobiose, and also active for xylotriose and xylo-oligosaccharide, gentiobiose, and cellobiose; feruloyl esterase acts on the ferulic acid ester of the arabinose in normal circumstances, in addition, it also acts on coumaric acid ester in some cases. Owing to the high heterogeneity structure of hemicellulose, different sugar molecules conformation numbers, nonsugar components, and the complicated connection bonds, this article does not conduct an in-depth discussion of specific enzymes. Instead, we list the basic activity, degradation effects on the biomass stocks, and the potential use of the hemicellulose enzymes (Merle, 2010).
4.1.2.1. Depolymerase
4.1.2.1.1. Xylanase
Xylan synthesis from different plants depends on poly β-(1–4)-xylopyranosyl backbone, and depolymerase acts on β-(1–4) or β-(1–3) bonds between xylans. Under the effect of these two enzymes, xylanase anomeric carbon configuration of the reducing end is retained. Most enzymes belong to two different categories of structural xylanase glycosyl hydrolase family: GH10 owns high molecular weight/low isoelectric point, and GH11 has low molecular weight/high isoelectric point. The former has a wider range of catalytic functions, and higher efficiency on the hydrolysis of xylan with a high degree of substitition. It is reported that xylanase has a cellulose-binding domain. Most xylanase is characterized by identifying the types of xylan hydrolysis and the final product series. The major products of most endoxylanase are xylobiose and xylotriose. Most endoxylenes can hydrolyze unsubstituted xylose and the hydrolysis of xylan side chains mainly relies on special xylanase.
Exoxylanase (3.2.1.37, 3.2.1.72, 3.2.1.56) usually has a hydrolysis specificity. β-Xylanase (EC 3.2.1.37) is more efficient on the high xylan. In addition, EC 3.2.1.72 is used for β-(1–3)-connected xyloside in most cases and exhibits limited degraded activity. Comparing with the endo xylanase, the enzyme protein of exoxylanase is usually larger (the molecular weight is larger than 100 kDa) and composed by two or more subunits. Due to the low content, little exoxylanase has been identified (Yang, Yao, & Fan, 2005).
4.1.2.1.2. Mannanase
Endomannanase randomly hydrolyzes the mannan and β-d-1,4 mannoside bonds, such as glucomannan, galactomannan, and glucomannan. Compared with xylanase, only a few of mannanase have been identified. Mannanase from T. reesei (a kind of bacteria) has a similar mutidomain structure of cellulose hydrolysis enzymes. The most common hydrolysis products of galactomannoza and glucomannan are mannobiose, manninotriose, and an oligosaccharides mixture. Hydrolysis products depend on the substitution degree and distribution of bonds.
Endonucleases release oligomer mannose and the further degradation requires β-mannosidase (1,4-β-d-mannanase, EC 3.2.1.25) and β-glucosidase (EC 3.2.1.21). β-Mannosidase and β-glucosidase catalyze the hydrolysis of oligomeric mannose and remove mannose or glucose from the nonreducing ends continuously. β-Xylosidase from T. reesei and β-mannosidase from Aspergillus niger can hydrolyze xylan and mannan, respectively, and release them by continuous exocutting effects (Zhao, Xue, & Ma, 2009).
4.1.2.1.3. β-Glucanase
Endoglucanase, which is often considered to be a cellulase family member, has a higher affinity for cellulose and also acts on xylan and mixed β-(1–3,1–4)-glucan. Just like endoxylanase, β-glucosidase can break the internal β-(1–4) or β-(1–3) bonds within glucose chains, and produce a reducing and a nonreducing end. EC 3.2.1.4 is one of the most important β-glucosidase; the β-d-(1,3) endoglucanase (EC 3.2.1.39) can act on the β-(1–4) glycosidic bond and its activity is limited on interconnected β-glucan. Endo-1,3 (4)-β-glucanase (EC 3.2.1.6) also has an activity of endoglycosyl hydrolase.
For cellulases and endoglucanases, a lot of researches have been carried out on fungus T. reesei. Such fungi produce a variety of cellulase, which act on cellulose synergistically. In T. reesei, Cel7Bis a major endoglucanase, accounting for 6–10% of total T. reesei cellulose. They have broad acitivity toward solid and soluble substrate, such as CMC, xylan, and glucomannan. Furthermore, endoglucanase Cel5A is also active toward solid and soluble substrate (CMC, mannan), but cannot act on xylan. These enzymes account for 10% of the total T. reesei cellulose. Little endoglucanase (Cel2A, Cel45A) has been reported with a variety of specific activity and can hydrolyze solid or soluble substrates (Li, 2004).
4.1.2.1.4. Xyloglucanase
Xyloglucanase is the main hemicellulose polymer during the growth of primary cell wall in plants and is hard to distinguish from cellulose and xylan. Xyloglucan is closely linked with cellulose microfibril though hydrogen bonds, providing loading network for cytoderm and protecting cell wall from disintegration under osmotic pressure. Nowadays, a lot of researches focus on finding key enzymes with controlling and modification effects during the cell walls amplification process. Though the branching degree of xylan is not complicated, xylose on xylan and other substituents makes enzyme digestion of xylan more difficult than cellulose and β-glucan. Xylose can form side chains with d-pyran galactose and l-fucose, but rarely with l-arabfuranose. It is reported that the enzymes, which can digest plant cell walls such as endoglucanase, xylan-endoglycosyltransferase, and exoglycosidase (taking α-fucosidase, β-galactosidase, for example), can digest xyloglucan. Parts of cellulose can also digest xyloglucan skeleton. Endoglucanases including xyloglucanase and xyloglucan specificity form a new kind of polysaccharide degrading enzymes, which can attack the main carbon chain, even replacing the anhydroglucose unit. Some glucanase only acts on specific xylose replacement type, and other enzymes have a broader substrate scope. A xyloglucan from A. niger can work on a variety of β-glucan, and shows the highest activity toward Tamarindus xyloglucan. Now a new kind of enzyme derived from plant is found to modify xyloglucan through endocuting, hydrolysis, and glycosyltransferase effects (Merle, 2010).
4.1.2.2. Debranching Enzyme
The glycoside lateral groups connected by xylan and glucomannan are mainly wiped off by α-glucuronidase, α-arabinfuranosidease, and α-d-galactosidase. The acetyl groups and hydroxycinnamic acid substituent group connected to xylan are removed by acetyl xylan esterases and ferulic acid/p-coumaric acid esterases. Usually substrate exhibits a feedback inhibition effect on the enzymes. There are significant differences among side debranching enzymes: some of them only hydrolyzed short chain oligosaccharides, which are mainly produced by backbone depolymerization endonuclease; others were able to remove branches on the whole polymer. But the latter is more suitable for the majority of oligomeric substrates. In the presence of auxiliary enzymes, synergy effects between the hemicellulases components can promote endoxylanase activity.
4.1.2.2.1. α-Glucuronidase
α-Glucuronidase (3.2.1.139) catalyze hydrolysis of xylan into glucuronic acid or 4-O-methyl-glucuronic acid. It is reported that the enzyme substrate can be long chain xylan, and also can be the substrate with only one xylose at the nonreducing end. A membrane-bound bacterial enzyme only act on soluble oligosaccharide derived from xylan. For wheat xylan substrate, the synergistic action of α-glucuronidase and endoxylanase produces highly dissociative 4-O-methyl-glucuronic acid (Pei, Yemin, & Azure, 2003b).
4.1.2.2.2. α-Arabinosidase
α-Arabinosidase can cleave arabinose side chains from the xylan backbone, which synergizes with ferulic acid and acetyl xylanase.
4.1.2.2.3. α-d-Galactosidase (Pei, Yemin, & Azure, 2003a)
α-d-Galactosidase mainly acts on the α-galactose side chain on the 0–6 positions of mannose skeleton units and can hydrolyze mannosanin wood, especially for galactomannan and galactoglucomannan. Although this enzyme is important for coniferous wood pulp, the corresponding researches are very limited (Li, 2004; Yang & Lee, 2006).
4.1.2.2.4. Acetyl Xylan Esterase
The acetyl group is present in many of the hemicellulose and has the highest number in glucomannan and galactomannan. Cereals and broadleaf wood have higher levels than coniferous wood in acetylation of xylan. Acetylation reactions in coniferous wood mostly occurs in glucomannan. The primary role of acetylation is to maintain the solubility and hydrability of hemicellulose. The removal of acetyl group from xylanand galactomannan will cause a sharp decline in solubility for polymers. The release of acetyl group from the main chain will affect microbial growth (e.g., reduced pH). The release of acetyl group inhibits the growth of microorganisms, which is a considerable problem in the process of biomass conversion (Chen Yan, 2010).
4.1.2.2.5. Ferulic Acid Esterase
Ferulic acid mainly presents in cereal, broadleaf wood, and pectin. In the xylan, they connected by an ester bond to the 2C of the side chains of arabinose backbone. The main function is to provide a three-dimensional stable structure for polymerization. Ferulic acid esterase (FAEs) can act on ferulic acid and coumaric acid. Some FAEs can react with polymer or xylo oligosaccharides. FAEs also act on xylan and pectin (Ceng Wei, 2009).
4.1.2.3. Hemicellulase for Different Biomass Feedstocks
Xylan composition varies with the type of substrate, and shows the biggest difference among coniferous wood, broadleaf wood, and herbs. Debranching enzymes include arabinosidase, FAE, coumaric acid esterase, and xylosidase. Furthermore, the removal of synergy effect between the side chain and the polymer backbone can enhance degradation rate of endoxylanase. Therefore, xylan treated by a mixture of depolymerase and debranching enzymes is much easier to degrade than those treated by acetyl-xylanase (Poutanen & Sundberg, 1988; Saha, 2000).
Xylan in coniferous wood is mainly composed of β-(1–4)-d-xylopyranose and the side chain is rich in 4-O-methyl-glucuronic acid. The ratio reported between uronic acid and xylose depends on the species and extraction methods, which ranges from (2:10) to (7:10). α-l-arabinofuranosyl units are linked to the backbone by 1,3-glycosidicbond. Unlike the xylan in hardwood and herbs, xylanin coniferous wood does not contain acetyl xylan groups. Therefore, the corresponding debranching enzyme does not contain acetyl esterase, which is mainly composed of α-glucosaccharase and α-l-arabinofuranosidase (Puls, 1997).
Xylan in broad-leaved wood is mainly composed of β-d-xylopyranose units, which also may contain methyl-4-O-α-d-glucuronic acid and acetyl side chains. 4-O-methyl glucuronic acid is linked to xylose backbone through O-(1–2)-glycosidic bond, an ester bond mainly in the C-2/C-3 hydroxy group. For broadleaf trees and herbs, the debranching enzymes mainly include acetyl esterase, arabinofuranosidase, and FAE. For softwood, it requires a higher α-glucosidase activity.
In addition, the specificity of depolymerase for side chain and structure is often related to the residues in the side chain. Endoxylanase randomly cuts 1,4-β-d-xyloside within the backbone and exhibits high specificity toward the type of sugar, the chemical bond, and the substituent.
Xylan hydrolysis is the dynamic equilibrium of three processes (removal of the side chain groups from the polymer backbone, reducing the length of the chain, and hydrolyzing the oligosaccharides into free monosaccharides. To achieve the best enzymatic efficiency, the enzymes need to work synergistically.
Glucomannan and galactose–glucomannan (GCM) are the major hemicellulose in coniferous wood. The structure of GCM changes with the species and cell wall positions, and is usually linked to the backbone made of β-(1–4)-d-mannopyranose and β-(1–4)-d-glucopyranose. Endomannanase and β-glucan are the main backbone depolymerase. β-Mannosidase and β-glucosidase only work in the hydrolysis of oligosaccharides (Puls, 1997).
The minority of hemicellulose in biomass include arabinogalactan, xyloglucan, and β-glucan. Arabinogalactan mainly exists in coniferous wood composed by linear β-(1–3)-d-galactopyranose. Although β-(1–4)-arabinogalactan is also found, it is mainly in the highly substituted C-6 position. As a debranching enzyme, β-glucuronidase can assist the polymerization of β-galactan. Xyloglucanase found to degrade the β-(1-4)-d-glucopyranose skeleton were mainly from the GH74 family. Among the enzymes, α-xylosidase has the widest range of applications (Tenkanen, 1998).
In addition, different preprocessing methods caused a different impact on the chemical structure of the hemicellulose, thus leading to different requirement for different types of enzymes: for example, the hydrolysis of polysaccharide in birchneeds xylanase, β-enzymes, and acetyl xylan esterase. The only usage of xylanase can only get 10% of the product. So, the selection of degrading enzymes must be based on the structural features of substrate.
4.1.3. Lignin-Degradating Enzymes
Lignin-degradating enzyme systems are very complex. In recent years, many scholars have carried out studies on the catalytic mechanism of lignin decomposition. These enzyme systems include extracellular peroxidase, 1ignin peroxidase (LiP), manganese peroxidase, (MnP), and extracellular phenol oxidase-laccase (laccase, LaC). In addition, aryl-alcohol oxidase (AAO), gyoxaloxidase (GLOX), glucose oxidase (gucose-l-oxidase), phenol oxidase, and so on took part in or have some impact on degradation process of lignin. Furthermore, the bacteria can produce two new enzymes: FAE and coumaric acid esterase, which act on lignocellulosic material and produce ferulic acid and p-coumaric acid. These two enzymes work synergistically with xylanase for the decomposition of hemicellulose lignin polymer without the production of mineralized lignin (Howard, Abotsi, & Jansen van Rensburg, 2003).
4.1.3.1. Lignin Peroxidase and Manganese Peroxidase
Both LiP and MnP are extracellular iron-containing heme proteins with glycosyl, are known as heme peroxidase. LiP is a glycoprotein, consisting of 10 long protein chains and a short protein chain, which is similar to other peroxidases, running a typical catalytic cycle; MnP is a kind of glycoprotein, which were also composed of ten long protein chains and a short protein chain.
The activity center of LiP consists of a heme group together with two Ca2+ for stabilizing of the structure. The activity center of MnP is basically the same as LiP, but has two more Mn2+. The main difference between the two is that carbon ends of LiP are between two propionates in heme, but carbon ends of MnP are separated from the heme. In addition, MnP owns five disulfide bonds but only for LiP. In the catalytic process of lignin degradation, LiP and MnP capture an electron from the phenolor a nonphenolic compound, changing the latter into a cation group and leading the bond cleavage of the lignin. LiP mainly oxidates phenol into phenoxy oxidation residues; MnP also mainly oxidates phenol into phenoxy oxidation residues, but the concrete roles of the enzymes have not been identified.
4.1.3.2. Laccase (LaC)
Laccase, a copper-containing polyphenol oxidase, can be divided into two categories: rhus laccase and fungal laccases. Those enzymes are mainly from lacquer and fungi. Due to high sugar content in fungal laccases, it was not until 1998 that humans obtained the first laccase crystal from Coprinus cinereus. Laccase can directly use O2 as a substrate without the presence of H2O2 and other second substrates. In this process, laccase itself loses the electrons or protons from hydroxy and transforms into free radicals. Laccase can also transform lignin into phenoxy radicals, which can further take part in copolymerization or homopolymerization reactions. These properties have made laccase gain a growing concern in the comprehensive utilization of lignin.
In addition to LiP, MnP, and LaC, there are also glucose oxidases, glyoxal oxidases, mellow oxidases, and catalase involved in the lignin degradation process. But so far, the specific role of each enzyme is not entirely clear in lignin degradation (Chi & Yin, 2007; Puls, 1997).
4.1.4. Cellulase Cofactors
For a long time, the study of cellulase has mainly focused on glycoside hydrolase. Despite a lot of exploration, there has still been no solution to the problem of the degradation of the natural crystalline cellulose due to ignorance of the role of supermolecular structure of cellulase in the enzymolysis process. Researches from the perspective of matter and energy found that the dense structure formed by hydrogen bonds is the bottleneck in enzymatic of cellulose.
The first stage of cellulose hydrolysis is the separation of oligomerization from the solid cellulose surface, which is the rate-limiting step of the hydrolysis. In the cellulose hydrolysis process, in addition to the synergy between the three cellulases, there are participation of other proteins. These proteins, which can promote the hydrolysis process, are called the cofactor of cellulase (Lu, Chen, & Ma, 2006).
Reese put forward the existence of a hydrogen enzyme in 1950, which could destroy the crystal structure of cellulase and enhance the accessibility of cellulose. Cosgove discovered the expansion protein in plants, which is considered to be most likely to play the role of “hydrogen enzyme.” In the process of plant growth, expansion protein can induce the wall to loosen and expand in a reversible manner. This process causes the sliding of polymers by breaking the hydrogen bonds within the polymer network. Expanding protein itself does not control glycoside hydrolase activity, but can weaken the strength of the filter paper, showing synergistic effects with cellulase in the hydrolysis of filter paper. Saloheimo found Swollenin protein owning the similar gene sequences with plant expansion proteins. In the hydrolysis of filter paper and cotton experiments, Swollenin proteins can destroy the structure of the cellulosic substrate, without reducing sugar (Saloheimo, Paloheimo, & Hakola, 2002; Levasseur, Saloheimo, & Navarro, 2006). Shandong University in China also conducted fruitful researches on Swollenin protein, including gene clone, protein structure analysis, and heterologous expression (Yao, 2007).
In addition, researchers also found a number of cofactors of nonhydrolysis cellulase. Gao Peiji et al. (Liu, Fan, & Sushi, 2008) separated a protein, which could weaken the absorption strength of cotton cellulose in hydrogen zone, lead to the expansion of cotton cellulose and chitin, and produce no reducing sugars. All these results matched the characteristics of Swollenin protein. Qiu applied laccase in the promotion of enzymatic digestibility for lignocellulose substrate. With the synergistic effect between laccase and cellulase, the reducing sugar content was increased by 37.9% and the ethanol production was increased by 13.8%. Analysis found that lignin got partly degraded after laccase treatment. This kind of ring-opening reaction transforms the substrate surface into a net structure, so that the accessibility of cellulase to the substrate has been enhanced significantly (Qiu & Chen, 2008).
4.2. The Setup of Biomass Invertase Platform
4.2.1. The Enzyme Production
Since bacterial α-amylases could be produced from scale fermentation, fermentation has became the main method for enzyme production. Depending on the cultivation modes, fermentation can be divided into submerged fermentation, solid-state fermentation, and immobilized cell culture. At present, the prevailing way is the submerged fermentation. Compared with traditional enzymes extracted craft from plant, the fermentation method has advantages in short production cycle, high enzyme yield, and less impact on the environment. However, it involves high requirements on the equipment and production craft (Chen, 2008). This section mainly focuses on fermentation technology for microbial enzymes.
4.2.1.1. Preparation of Cellulase Production
1. Producing strains
Bacteria, fungi, and actinomycetes can produce cellulase, among which fungi Trichoderma has been widely studied and applied due to the high production ability of cellulase and high enzyme activity. The most researched fungi include T. reesei, Trichoderma koningii, and Trichoderma viride. In addition to Trichoderma, Actinomycetes are also the ideal strain due to their simple structure, which is easy for genetic analysis. A lot of researchers are committing to Actinomycetes enzyme production research. Mutagenesis is the main source of excellent fermentation bacteria for cellulase. Taking Trichoderma TH (Trichoderma pseudokon-ingii) as the starting strain, mutant UVIII obtained by UV could tolerate high concentrations of glucose, and owned 100-fold higher sensitivity to the inducer than the parent strain. At the meantime, the glucose absorption capacity declined significantly, making the bacteria partially remove the repression of glucose inhibition. With the development of modern molecular biology, genetic engineering bacteria has also become a source of cellulose fermentation bacteria (Wei & Li, 2008; Zhang, Liu, & Gao, 2009).
2. The fermentation medium
Cellulase is a kind of inducible enzyme, whose biosynthesis was regulated by inducer. The fermentation medium usually choose the splintered, pretreated, and plant fiber contained raw, waste paper, vinasse as the main inducers and major carbon source together with nitrogen source and suitable inorganic salts. Research shows that adding 15% wheat bran, 2% urea, 1% ammonium sulfate, and 0.15% of KH2PO4 in brewers’ grains can improve enzyme activity and protein content significantly in the product. For liquid fermentation, these materials are made into liquid medium supplemented with excess water. In most researches, dry materials accounted for 3% of the amount of liquid medium. Xiaobin Yu et al. optimized liquid fermentation medium of Reesei cellulase WX2112 using response surface method and determined the best combination: 3.18% of soybean meal, 2.95% of wheat bran, 0.25% of KH2PO4, and 3.79% of avicel. The filter paper activity could reach a maximum of 10.53 IU/mL.
3. Brief introduction to the fermentation process
a. Solid state fermentation
Solid-state fermentation method could use straw powder and waste paper as raw materials. The materials are always mixed with koji, loaded under the curtain, or spread into a thin layer (thickness 1 cm), and ferments at a certain temperature in the culture room and humidity (RH 90∼100%). Its main feature is that there is no free water in the fermentation system, and microbes grow in the solid substrate with sufficient humidity and the fermentation environment is close to the natural state of microbial growth habit. It has many advantages such as more kinds of enzymes being produced, conducive to the degradation of natural cellulose, low investment, low energy consumption, high output, ease of operation, high recovery rate, no foam, and less environmental pollution. However, solid-state fermentation is susceptible to bacterial contamination; it is thus difficult to separate and purify the cellulase production, and tough to remove the pigment (Chen & Xujian, 2008).
The temperature is the primary factor for solid-state fermentation. Optimization of media and culture conditions are an important method to reduce enzyme costs, increase enzyme activity, and realize the industrial production. Generally, it is considered best to adjust the starting pH of the medium to acidic range, which will help the growth of fungi and inhibit the bacteria breeding in solid-state fermentation. Hongzhang Chen and coworkers (Xu, Chen, & Li, 2002a; Xu, Chen, & Shao, 2002b) proposed a novel fermentation model named gas double-dynamic solid-state fermentation: under the optimum pressure pulse range, pulse frequency, and the circulation gas rate, fermentation temperature can get better controlled and the maximum temperature gradient is 0.12°C/cm; water activity get well maintained within steam-exploded straw as fermentation substrate; the fermentation period of dynamic (60 h) is a third shorter than the static fermentation period (84 h) and the enzyme activity increased by onefold from 10.182 to 20.36 IU/g. Under the pulsation pressure, microbial grows uniformly in the solid material layer, while there is no cell growth in the middle layer of the material under static solid-state fermentation condition. Gas double-dynamic solid-state fermentation lays the foundation for large-scale production of cellulase.
b. Liquid fermentation method
Liquid submerged fermentation is also known as full fermentation. In this method, straw and other raw materials are crushed and sent to a closed fermenter with stirring paddle and ventilation systems, cultivated with strain, and then sufficiently stirred with sterile air or self-priming the airflow. Gas and liquid can get as far as possible to contact to help the fermentation. Its main features are easy-to-control culture conditions, the avoidance of infection, and high production efficiency. As one of modern biotechnologies, submerged fermentation has become an important research and technology both domestically and internationally.
Submerged fermentation generally uses a closed fermenter with a stirring paddle and ventilation system. The whole process from the medium sterile, medium cooling to fermentation are carried out in the same fermenter. Most fermentation time is about 70 h, and the temperature is generally less than 60°C. Inoculation quantity in liquid fermentation [generally 2∼10% (v/v)] is significantly lower than solid state fermentation. Zhang Dongyan studied the suitable fermentation conditions of Viride A S1313711, Trichoderma A S1312774, Trichoderma A S1313032, and Trichoderma ACC131167 for enzyme production. The results showed that the optimum temperature is 28°C, and appropriate initial pH for enzyme production is 4.5–5.5. The optimal parameters of Gibberella fujikuroi for cellulase production are as following: inoculation size is 5%, the incubation time is 120 h, the culture temperature is 28∼37°C, and initial pH value is 5–6 (Zeng & Wang, 2009).
4.2.1.2. Hemicellulase Production
Studies on xylanase is primarily from fungi and bacteria. In most cases, the best activity is achieved at or near the temperature (about 40–60°C), neutral conditions (mainly bacteria xylanase) or slightly acidic conditions. However, some xylanase also been reported to have activity under extreme pH and temperature. In fact, it has been reported that xylanase remains active at a temperature of 5–105°C within pH 2.0–11.0. Some studies show that xylanase remained active when the concentration of NaCl up to 30%. These enzymes are ways that microorganisms adapt to extreme environments. Among these enzymes, thermophilic, basophils, and eosinophils xylanase been extensively studied. Conversely, the study on cold-adapted xylanase is much rarer (Shi, 2011).
Xylanase derived from fungi has higher enzyme activity, such as T. reesei rut, C-30,851 IU/mL; Fusarium oxysporum, 245 IU/mL; Thermoascus aurantiacus, 208 IU/mL; A. niger, 283 IU/g; and Melanocarpus albomyces, 9300 U/g. The enzyme activities are not strictly comparable because the detecting step and the reaction conditions are not the same, such as temperature, incubation time, and substrate used. The stability of xylanase from fungal reported in alkaline and high temperature environment is not good except for M. albomyces. This enzyme exhibits good stability and high activity at pH 10.0, 70°C in solid-state fermentation, whose half-life is about 2 h.
The strains producing alkali resistance, high temperature, and high xylanase activity include: Bacillus sp. Sam3, xylanase activity of 131 IU/mL, pH 7.0–9.0, 60–70°C; Bacillus sp. NCIM59, xylanase activity 500 IU/mL, pH 7.0–10.0, 60–70°C; Bacillus cirulans Ab16, xylanase activity 50 IU/mL, pH 5.0–9.0, 55–80°C, and C. absonum CFR-702, xylanase activity 420 IU/mL, pH 6. 0–9.0, 78–85°C (Quan & Zhao, 2010).
4.2.1.3. Preparation of Lignin-Degrading Enzyme
1. Producing strains
MnP is widely present in the white-rot fungus, such as the famous biopulping insect species, wax bacteria. It is easy to detect the presence of MnP when white-rot fungus lives on the lignocellulosic substrates. Almost all the Basidiomycetes, which cause wood rot and habitat in the soil degradation layer, can produce this enzyme. These Basidiomycetes mainly belong to wood decay fungi, including Corticiaceae, Stereaceae, Hericiaceae, Ganodermataceae, Hymenochaetaceae, Polyporaceae, Strophariaceae, Tricholomataceae, and so on. However, bacteria, yeasts, filamentous fungi, and mycorrhizal fungi do not produce MnP.
Laccase (EC 1.10.3.2) is present in a wide range of fungi strains including Basidiomycetes, Ascomycetes, and Deuteromycetes, and some insects, bacteria, and plants also produce laccase. Laccase owns monomer, dimer, and tetramer forms with monomer as the general form. Laccase from the white-rot fungus owns the following characteristics: molecular weight is between 60∼80 kDa, glycosylated, containing 15–20% carbohydrate, owning an acidic isoelectric point (the pH value is between 3.5 and 7.0). Laccase can be produced by most of the white-rot fungus stains, but except Phanerochaete chrysosporium, which is well-known, for the lack of laccase production ability.
The lignin peroxidase was first discovered in P. chrysosporium. At present, only a few white-rot fungi can produce LiP. The molecular weight of LiP is about 40 kDa, glycosylated, having an acidic isoelectric point. In addition, they contain a heme iron protoporphyrin IX (Chi & Yin, 2007).
2. Fermenting the substrate
External factors affect the production of lignin-degrading enzymes, which are carbon, nitrogen, oxygen, and trace elements. Among them, the fungus P. chrysosporium is the more researched is generally believed to produce LiP and MnP but no laccase. However, literatures reported that P. chrysosporium can produce a small amount of lignin peroxidase and laccase under severely controlled concentration of nitrogen.
Lignin-degrading enzymes are produced in an oxygen environment. When using glucose as a carbon source, the lignin-degrading enzyme synthesis was inhibited in P. chrysosporium. While using cellulose as a carbon source and cellulose is completely consumed and no oxygen is required, strains began to generate LiP. The right amount of manganese can improve LiP activity of P. chrysosporium. Mn is an element required in MnP synthesis; the amount of Cu2+ can promote the synthesis of strain laccase.
Inducers and surfactant have an important role in the synthesis of lignin degrading enzymes. The inducer studied most was resveratrol. Different strains have the corresponding optimum surface agents. Polyethylene can stimulate the synthesis of lignin peroxidase, and 3,4-dichlorophenol can maintain the activity of LiP and MnP at a high level over a period of time. In the fermentation process of P. chrysosporium, adding HgCl2 can improve stability and activity of peroxidase, especially LiP. When using ferulic acid (FA, 4-hydroxy-3-methoxycinnamicacid) as substrate, LiP will be out of activity. p-Coumaric acid and guaiacol can improve laccase production of white-rot fungus Trametes sp. I-62 significantly. 2,5-benzyl alcohol is the best laccase inducer, followed by 3,5-dimethoxybenzyl, and 3,4-benzyl alcohol was the worst laccase inducer (Hui, 2006).
Chen Hongzhang used steam-exploded wheat straw as a substrate for lignin enzyme production through solid-state fermentation. Cellulosic feedstocks, especially grasses, have been partially degraded after steam explosion treatment and the degradation products can replace expensive resveratrol as lignin gene expression inducer. Substrate produced from lignin peroxidase is rich in lignin. Cellulose-rich substrate is conducive to the synthesis of laccase (Lu, Zhang, & Hong, 2005).
3. The fermentation process introduction
The fermentation process of microbial enzyme production is closely related with environmental factors. Fermentation methods have a great influence on the laccase production of white-rot fungi. According to the training methods, it can be divided into two categories: liquid fermentation and solid fermentation.
4.2.1.3.1. Liquid Fermentation
Liquid fermentation refers to the process of mycelial growth and enzyme production in liquid medium containing certain nutrients. Since this method can facilitate industrial production and application, therefore, the reports of enzymes production in white-rot fungi through liquid are relatively common. Most studies have shown that adding an appropriate amount of inducing agent or Cu2+ in liquid medium can promote laccase production from white-rot fungus significantly.
In addition, some metal elements, such as copper, manganese, iron, zinc, and so on, are an integral part or enzyme reactive group, which cannot be ignored. Many studies (Birhanli & Yesilada, 2006; Lorenzo, Moldes, & Sanromán, 2006; Rosales, Couto, & Sanromán, 2007; Gassara, Brar, & Tyagi, 2010) have confirmed that adding a certain amount of Cu2+ in the medium can promote laccase production of white-rot fungus significantly.
4.2.1.3.2. Solid-state Fermentation
Solid-state fermentation is a method that uses solid medium to culture mycelial and harvest enzymes on the substrate surface in the absence of or containing a small amount of liquid. With the continuous development and in-depth study, fungal enzyme production by solid fermentation has attracted more and more scholars’ attention. Research and practice show that solid-state fermentation of white-rot fungi can get higher laccase production compared to the liquid fermentation. Solid fermentation supplies more oxygen circulation, which is conducive to the growth of microorganisms. Furthermore, the process is static, so the energy consumption of mechanical power can be omitted, and the subsequent process is relatively simple. These methods are close to the wild state, in particularly suitable for enzyme production.
Scale expansion of white rot fungi solid-state fermentation (SSF) has also obtained more and more attention from researchers. Mass transfer system laid great impact on laccase yield. When using barley bran as noninert substrates, disc reactor is more suitable for white-rot fungus Trametes versicolor for laccase production (Couto, Sanromán, & Hofer, 2004; Couto, Moldes, & Liébanas, 2003). The maximum laccase activity reached 3.5 IU/mL, six times those in the other two fermenters. When using orange peel as a substrate in solid-state fermentation, the disc reactor is also applicable to white-rot fungus Trametes hirsuta for laccase production with the highest activity as 12 IU/mL. Because the disc reactor is better than the other two solid-state fermenters, scholars believe that the mass transfer would produce a certain shear force in the submerged and expanded bed reactors, affecting cell growth and enzymes biosynthesis. Therefore, design and mass transfer system is a key factor affecting the efficiency of enzyme production.
At present, reports about the application of solid fermentation for laccase production from the white-rot strain are relatively rare. This is mainly because the solid-state fermentation tank is facing some insurmountable problems. In addition to equipment design and mass transfer system, fermentation process parameters include pH, temperature, aeration, and oxygen delivery, and humidity, which are difficult to control. Therefore, the current study focused on the improvement of existing solid-state fermentation systems or design of novel fermentation equipment, such as new RITA (Recipient Immersion Temporaire Automatique) batch immersion systems (Boehmer, Suhardi, & Bley, 2006). RITA immerses mycelium growing on the solid substrate into liquid medium. The advantages of this reactor are that it not only avoids the mechanical shearing effect on bacteria, but also is able to overcome the metabolites and organic dyes for cell growth and enzyme inhibition.
Another way for efficient production of laccase from white-rot fungi is to seek a suitable carrier for the heterologous gene expression. The presence of glycosylin laccase can promote the hydrolysis of laccase protein, compared to other industrial production oxidoreductases; expression of white-rot fungi laccase in the heterologous host is very difficult. Therefore, the present study on laccase production is mainly concentrated on the optimization of fermentation conditions.
In recent years, from the study of laccase production from white-rot fungi, either liquid or solid fermentation, a new feature has emerged that the natural substances is being used, especially the use of lignocellulosic organic by-products generated in industrial and agricultural production, as the culture substrate. These organics include wheat bran, bagasse, rice straw, wheat straw, wood chips, straw skin, lees, soybean meal, corn cob, deinking sludge, vines, and olive mill wastewater. Most of them are usually rich in carbohydrates, able to provide nutrients for bacteria growth and metabolism. Furthermore, these organics also contain a certain amount of lignin, cellulose, and hemicellulose, regarded as an inducer of laccase production in white-rot fungi. It is reported that wheat bran also contains a lacquer enzyme-inducing substances-ferulic acid (approximately 0.4–1.0%) with a strong inducing effect on laccase production. Although so far there are no reports on other natural substrates containing special inducing substances, the feasibility of these natural organic matters as fermentation substrate for white-rot fungus has been widely recognized.
4.2.1.4. Cutinase Production
With nitrophenyl butyrate as the screening substrate, a Thermophilic bacterium strain producing cutinase was screened. Under the suitable culture conditions, cutinase activity reached 19.8 U/mL. By pNPB hydrolase activity detection, pure natural cutinase was separated from fermentation supernatant of Thermophilic sp. (T. fusca) by sequential treatment of ammonium sulfate precipitation, hydrophobic interaction chromatography and anion-exchange chromatography separation. Thermophilic sp. cutinase has good thermal stability and pH stability. The optimum temperature is 60°C and the optimum pH for the reaction is 8.0, which is in line with the required clean production characteristics of the textile industry (Li, Liu, & Chen, 2009).
4.2.2. Chemical Modification Enzymes
Chemical modification aims at changing the properties of natural enzymes for excellent characteristics and wider applications. Generally, the conformational change may occur following the chemical modification of the native enzyme and improve the enzyme activity and stability. We can say that the enzyme chemically modification theory provides an experimental basis and proof for the relationships between the structure and function of biological macromolecules. Chemical modification is very effective to improve the enzymatic properties and enlarge its application range.
Experimental studies have shown that the appropriate chemical modifier and modification conditions can not only retain the enzymatic activity, but also can improve enzyme stability to heat, acids, alkalis, and organic solvents and change enzyme substrate and the optimum pH. There are a lot of enzyme chemical modification methods, but the basic principle is the use of chemical characteristics that the modifier possessed, directly or indirectly, reacting with amino acid residues in enzymes molecules, thus transforming into the enzyme molecule.
When the enzyme is chemically modified, the following must be noted: (1) after the enzyme treatment by pH, oxidation, or reduction, the total amino acid analysis should be carried out, in particular, those are difficult to analyze (such as tryptophan, methionine); (2) because the modifying agent selected for the model compounds or enzymes do not necessarily apply to others, therefore one should find out whether other enzymes have been modified; (3) controlling the modification reaction conditions, such as buffer components; (4) identify whether there is a significant difference between optimum pH of the natural enzyme and optimum pH of enzyme after modification.
Until now a variety of types of protein modification agents has been developed, including: (1) small molecules, such as acetyl imidazole, haloacetic acid, N-ethylmaleimide, carbodiimides, coke, diethyl tetranitro methane; modifying agent; and (2) macromolecules such as polyethylene glycol (PEG), polyamino acids, acetic acid/propionic acid copolymer, carboxymethyl cellulose, polyvinyl pyrrolidone, dextran, and cyclodextrin. Whether small molecule or macromolecular modifying agent, they can chemically react with a specific amino acid residue to form a covalent bond.
4.2.2.1. Chemically Modified Cellulases
Maleic anhydride is often used as a chemical modifier for preparing alkali cellulose using acidic cellulase as substrate. Maleic anhydrides electively modified lysine group of cellulase, which extended cellulose tolerance to pH from 8.5 to 10.0. Using 0.5 mol/L maleic anhydride under reaction conditions pH 8.0, 2∼4°C with cellulase 25 min, the half-life of the modified enzyme under pH 8.0, 9.0, 10.0, and 11.0 was 13.08, 121.58, 126.0, and 187.30 h, respectively. Therefore, enzyme stability is greatly improved. It is reported that the modified maleic anhydride and N–bromosuccinimide cellulases are also good modifier, and the half-life of the modified enzyme at 30, 50, and 85°C are up to 120 min. The performance of the resulting cellulase was also superior to the original detergent enzymes.
The cellulase modified by PEG owns significant enhancement on the thermal stability. Therefore, the capacity to resist heat also increased. At the same time, the affinity between the modified enzyme and the substrate also become higher than the natural enzyme (Li, 2007b).
4.2.2.2. Chemical Modification of Laccase
With deep understanding on the structure and function of laccase, it was found that lysine (lys) residues is not the active center of laccase, which provides a theoretical basis for the chemical modification of laccase. At present, PEG, dextran, phthalic anhydride (PA), and citraconic anhydride (CA) have been applied to laccase chemical modification, and the effects of modification were also studied.
Hua et al. modified laccase with PA in order to improve its stability against high temperature and acidic pH. The modification ratio of amino lysine groups of laccase is determined as 63.8% by TNBS method. It is shown that the modified laccase maintained the secondary and tertiary structure of its native counterpart but has a higher affinity to ABTS and a significantly improved thermal stability, as indicated by the extension of the half-life of enzyme activity at 55°C from 192.5 to 532.4 min. The suitable pH range is also expanded from pH 5.9∼7.8 to pH 4.5∼8.4. The degradation ratio of anthracene is increased twofold, making the modified laccase promise for the removal of PAHs.
Native laccase was applied widely in industry. It was becoming very important and urgent because native laccase was amenable to activity loss under its application conditions. A commercial native laccase named DeniLite IIS with wide application has been researched. This laccase was modified with different anhydrides, such as phthalic anhydride (PA), succinic anhydride (SA), and maleicanhydride (MA). SA was confirmed as a best modification reagent for laccase according to enzymatic; activity and stability enhancement of modified laccase. The influences of three factors of phosphate buffer pH and SA concentration and modification time on enzymatic activity and stability were studied by L9 (34) orthogonal experiment. The results showed that no interaction existed between these three factors. The optimal conditions for modification were that phosphate buffer pH was 7.5, SA concentration was 2 mmol/L and modification time was 1.0 h. SA modification of laccase under the optimal conditions caused enzymatic activity increased by 50% and thermal stability at 50°C for 30 min enhanced by 15%. This research offered a suitable method for property reform of native laccase (Ya-hong, Gao, & Zheng, 2011).
4.2.3. Enzyme Engineering
With the establishment and development of recombinant DNA technology, people remove dependence on natural enzymes in large part. Developments of genetic engineering enable people to obtain a variety of natural enzyme genes by cloning, and highly express them in heterologous microbial receptors, in the end massively producing them by fermentation technology. This method can greatly reduce the cost of the enzyme product, and also makes the production of rare enzymes easier.
4.2.3.1. Gene Cloning
Most of the earlier enzyme-cloning method was achieved by constructing is a library, such as DNA libraries and cDNA libraries. A DNA library takes genomes of the starting strain as research object, by random digestion of genome into varying sizes of fragments, random digestion fragments of varying sizes, and inserted them into the cloning vector. After positive clones were screened, the objective gene can be determined. A cDNA library is a combination of cloned cDNA (complementary DNA) fragments inserted into a collection of host cells, which together constitute some portion of the transcriptome of the organism. cDNA is produced from fully transcribed mRNA found in the nucleus and therefore contains only the expressed genes of an organism. Similarly, tissue-specific cDNA libraries can be produced. In eukaryotic cells the mature mRNA is already spliced, hence the cDNA produced lacks introns and can be readily expressed in a bacterial cell. While information in cDNA libraries is a powerful and useful tool since gene products are easily identified, the libraries lack information about enhancers, introns, and other regulatory elements found in a genomic DNA library.
With the fast development of network technology and gene synthesis technology, the current cloning method is more simple and straightforward. Cloning methods commonly used as following: (1) the synthetic method. According to the reported nucleotide sequence, researchers synthesized these genes in labs. (2) Specific primer amplification method. This refers to the method that designs the specific primer based on the nucleotide sequence and amplifies the similar gene from same species.
1. Cloning and expression cellulase gene
So far, more than 7,000 cellulose enzyme gene sequences and the corresponding amino acid sequences has been reported and the 3D structures of more than 500 celluloses have been published. These data are published in Gen Bank, EMBL, DDBJ, and so on. At the same time, almost all of the cloned cellulase genes have been expressed in E. coli (Li & Zhang, 2011). Nearly more than 100 cellulase and xylanase genes can be cloned and expressed in E. coli, mainly endoglucanase and β-glucanase. Researches on cellulase synthesis regulation, cellulose degradation mechanisms, and new enzyme molecule construction can be achieved by cloning methods. There are no operation challenges on DNA extraction, digestion, plasmid recombination, and transformation.
2. Cloning and expression xylanase genes
At the time of writing, more than 300 kinds of different xylanase genes sources have been reported, of which more than 100 kinds have been cloned and expressed in suitable hosts.
Endoxylanase genes derived from Penicillium owns 10 sequences, among which five belong to the Family 10, four belong to Family 11, and the other belongs to Family 7. Four inner endoxylanase genes have been reported from Penicillium funiculosum, three of which owns CBM domains. XynA gene encodes xylanase/cellobiohydrolase which is similar to family 7. XynB genes belong to Family 11 and xynD belong to Family 10. These enzymes include a catalytic domain, and a serine-aspartic acid-rich connecting region and a prediction CBM domain.
All of the xylanase-encoding genes described above have been expressed in Pichia yeast, E. coli, fungi, Aspergillus, Saccharomyces cerevisiae, and other expression systems. The optimum pH of xylanase derived from Penicillium is between 2.0 and 7.0, and the optimum temperature is between 40 and 60°C (Liu & Gao, 1998).
3. Cloning and expression of mannanase gene
Since Henrissat (1991) divided the mannanase into glycoside hydrolase Family 5 and Family 26, many researchers have studied the sequences of mannanase gene. Currently, there are hundreds of pieces of β-mannanase gene information in gene banks, most of which have been cloned in bacteria, fungi, and plants. Some mannanases can maintain its activity at extreme environments, such as the optimum pH of MAN5A P13 is 1.5 and optimum pH of MANN from Aspergillus sulphureus is 2.4. The production of Man 1 from Yarrowia lipolytica reached 5.9 g/L (1575 U/mL).
β-Mannanase gene from fungal is the most cloned and expressed. This gene can be isolated from Agaricus bisporus, Armillariella tabescens EJLY2098, Aspergillus fumigatus, P. chrysosporium, Aspergillus aculeatus, and so on. They have been expressed in Pichia yeast, S. cerevisiae, A. niger, and other heterologous expression systems (Cai, 2011).
4.2.3.2. Site-directed Mutagenesis of the Enzyme Molecule
When researching the natural enzyme and the mutant, it is usual first to obtain the information on molecular characteristics, space structure, and relationship between structure and functions. This information can be obtained through biochemistry, spectroscopy, and crystallography technologies. Then this information is used for enzyme molecular modification, which is called rational designment of enzyme molecular. Rational designment of enzyme molecular mainly includes chemical modification and site-specific mutagenesis. In contrast, the modification without enzyme information was called nonrational designment of enzyme molecular, which includes determinate evolution and hybrid evolutionary.
The foundation of rational designment is the scientific knowledge of protein construction for engineering prediction. When using this strategy, it is essential to obtain the information of protein structure, especially the catalytic site and the relationship between the structures and functions. In the typical case, the data efficiency of enzyme or homological protein determined the protein to modify. First, we should understand the role of the amino acid residues in the enzyme, then carry out the modification with site-specific mutagenesis, secondary structure elements, even the exchange of structural threshold. Finally, we can analyze the characteristics of mutant enzyme. In recent years, the rational designment strategy has been widely used in the research of relationship between enzyme function and the specific amino acid residue. Until now protein, engineering has been carried out on the cellulase from different glucoside hydrolase (GH) families, including GH5, GH6, GH7, GH8, GH9, and GH45.
1. Site-directed mutagenesis of endocellulase
Application of site-directed mutagenesis techniques on Thermobifida fusca and Bacillus N4 transformation can be changed the amino acid sequences within the cellulose enzyme, thus increasing the adapt range of pH, reducing product inhibition, and increasing enzyme activity.
In addition, the use of molecular biology techniques to build hybrid enzyme also made considerable progress. Studies on the extreme heat archaeal enzyme EGPh implied that the enzyme lacks a cellulose-binding domain, and E324 is the active center of the enzyme.
2. Site-directed mutagenesis of exoglucan cellobiohydrolase
The goal of rational design on endoglucanase is mainly to increase enzyme activity and stability (Zhang, Zhang, & Gao, 2006). Von Ossowski found that the outer ring from G245 to Y252 of Cel7 A of T. reesei have an impact on the enzyme activity. Wohlfahrt applied site-directed mutagenesis techniques onto T. reesei, replacing amide-carboxylic acids with carboxylic acid and making the mutant melting temperature changes. The modification improved the enzyme stability under alkaline conditions significantly. Though the catalytic rate on cellotetraose has not been alerted, the half-life of the two mutant enzymes under basic conditions increased by 2–4 times. Therefore, carboxylic acid can effectively change the pH-dependent properties in the protein engineering.
3. Site-directed mutagenesis of β-d-glucosidase
Compared to exoglucanase enzymes, the rational design on the endoglucanase was relatively less. Fukuda applied site-directed mutagenesis on G294 of on the Bgl I from Aspergillus oryzae, and obtained three mutants, G294, FG294, and WG294Y, whose enzyme activities were increased by 1.5, 1.5, and 1.6 times higher than the wild, respectively (Li et al., 2009).
4.2.3.3. Enzyme-directed Evolution
The relationship between protein structure and function is very complex, and the current understanding on the relationship is still very limited. When the knowledge of the molecular structure is very poor, the directed evolution become a powerful tool. Directed evolution can simulate the natural evolutionary mechanism without prior knowledge of the spatial structure of the protein. Its core is a random mutation plus selection, namely by PCR or other methods for gene random mutation or recombination, and make it expressed in the host cell, and then a high-throughput screen the mutant enzymes. For many cellulase enzymes, the three-dimensional structure has still not been resolved. Therefore, directed evolution has become a powerful tool for cellulase modification. Scholars have carried out a number of studies, and obtained a large number of mutant enzymes with improved activity, increased stability, or other properties (Ping, Liu, & Xue, 2009).
Thermoanaer-obacter tengcongensis MB4 endoglucanase Cel5A pretreated by error PCR was screened by the Congo red plate and obtained two enzymes with 135 and 193% folds activities of the wild enzyme. Endoglucanase (EngD) from mesophilic Clostridium cellulovorans recombined with CelE from Clostridium thermocellum and obtained a mutant enzyme with twofold increase on the enzyme activity at 50°C.
In the study of directed evolution of cellulase, researches on xonuclease are relatively less. This is mainly because the current exonuclease activity assay has not been unified, convenient, and fast, which restricts the throughput of mutations screening (Zhang et al., 2006; Li, 2007a).
4.2.3.4. Fusion Enzyme
Fusion enzyme refers to the fusion protein composed by two or more enzymes (Chen, 2008). A fusion enzyme belongs to hybrid enzymes. It combines the advantages of different enzymes and has obvious heterosis. Amino acids of fusion enzyme have differences in static electricity, acidity, acidity and ion; therefore, they have special characteristics that normal proteins do not have.
There are two ways to produce fusion enzymes: one is nonrational designment, mainly by building the library, and then screening out the fusion. Another method is rational designment, fusing the enzyme based on the key information of enzyme structure and functions. The fusion protein is usually conducted among homologs, because less similarity of the protein sequences increases the probability of fusion failure. The characteristics of the enzyme will change during the fusion treatment, including kinetic parameters, thermal stability, and optimum basic characteristics such as pH changes more easily.
Fusion enzyme construction strategy can be divided into three categories: secondary structure integration, function domain integration, and whole protein integration. It is a corollary to develop the fusion protein engineering technology. This technology is based on protein project and computer science, which makes the synthetic enzyme strategy possible.
4.3. Enzymes Conversion Platform for Biomass
4.3.1. Enzyme Synergy
Evolutionary selection pressure makes the microorganisms living on biomass use enzymes to degrade the biomass. Two collaborative system have been discovered: one is multicomponent cellulase secreted by fungus/bacteria and the other is cellulose secreted by anaerobic strains (Gruber, Kamm, & Kamm, 2007; Yinbo, 2011).
4.3.2. Multicomponent Enzyme System
There are two synergy forms between fungal cellulase (Mosier, Hall, & Ladisch, 1999): one is the synergy between exocellulase and endocellulase; and the other is the synergy between exocellulase and exocellulase (Goyal, Ghosh, & Eveleigh, 1991; Wood & McCrae, 1979).
Chanzy, Henrissat, and Vuong, (1984) proved CBHI enzyme can randomly bond to molecule chains of cellulose by studies of colloidal gold. Herissat et al. found that the ability of EGI for hydrolysis of microcrystalline cellulose is very low, and CBH I has no activity to CMC, but on microcrystalline cellulose. EG I and CBH I can synergistically degrade filter paper, microcrystalline cellulose, microcrystalline cellulose, and homogenized bacterial cellulose, but cannot degrade CMC synergistically. CBH I and EG I or EC II degrade insoluble cellulose synergistically, but the optimal ratio between the enzymes was different. Nisizawa found that CMC with a high degree of substitution has higher exonuclease activity than CMC with a low degree of substitution, suggesting that the low activity of CBH Ion CMC was related with substituent groups.
T. reesei is a representative fungal cellulase. As an efficient cellulose producer, it is capable of secreting a large amount of cellulase, including CBH I, or Cel7A (60%), CBH II, or Cel6A (15%), EG I or Cel7B, EG II, or Cel5A (20%), and other minor components (such as EG III or Cel3A, XynI or Xyn11A, and swollenin). The difference between CBH I and CBH II is whether can satisfy the degradation requirement of cellulose substrate.
4.3.3. Fiber Bodies
C. thermocellum is a typical hydrolyzing enzyme system of bacteria. This cellulase system consists of an approximately 200 KDa molecular, which has special structures including protein scaffold, anchoring threshold, cellulose binding threshold, and many cohesin domains. Molecular mass of fibrous bodies ranges from 50 to 5,000 Da and many fiber bodies also form polyfiber bodies through anchoring interactions. The catalytic domain of fiber bodies is similar to multicomponent fiber cellulase secreted by fungus. However, comparing with the multicomponent fiber cellulase, the hydrolysis efficiency of fiber bodies was much higher (Chen, 2011; Yinbo, 2011).
In summary, an efficient industrial cellulase enzyme system should include a variety of functional enzymes, and these enzymes must be able to perform synergistically.
4.3.4. Optimization of the CBH–EG–BG System
Since cellobiose is not only an inhibitor for CBH, but also is the precursor of fermentable sugars. Therefore, a complete cellulase system requires the use of BG to hydrolyze cellobiose. How to achieve mutual balance among CBH–EG–BG is the key issue to improve hydrolysis efficiency. Researches showed that T. reesei could secrete at least two kinds of BG enzymes under the abduction of cellulose. At the same time, comparing with the closed system, the filtration equipment, which gets rid of small molecules, can improve the efficiency of cellulose hydrolysis. Cellobiose is an inhibitor of CBH, the accumulation of which can decrease the hydrolysis rate (Gruber et al., 2007).
4.3.5. Design of the Multienzyme Complex
Studies have shown that the enzyme synergy can help to build a more efficient industrial enzyme system. However, there is a difference on the characteristics between enzymes. So there are still challenges for man-made enzyme synergy systems. We just make a brief introduction on the design of the multienzyme complex. This can be divided into four steps: (1) the selection of the enzymes; (2) the consistency evaluation among different enzymes; (3) determining of the mixing ratio among enzymes; and (4) the optimization of technological parameters.
Taking the hydrolysis of arabinose and xylose in wheat as an example, the minimal enzyme cocktail (MEC) includes four key enzymes. A slight difference on the substrate structures leads to different requirement on the compound proportion and amount of the enzymes used. The different compound enzymes and their hydrolysis results for different substrates are given in Table 4.1.
Table 4.1
Compound Enzymes and Their Enzymatic Hydrolysis Effects for Different Substrates
| Substrate | Abf II (%) | Abf III (%) | β-xyl | Xyl III (%) | Hydrolysis yield (mg.g DM) | |
| Arabinose | Xylose | |||||
| Water-soluble Arab xylan in wheat | 20 | 20 | 40 | 20 | 343.7 | 548.7 |
| Water-insoluble Arab xylan in wheat | 25 | 25 | 25 | 25 | 162.5 | 286.5 |
| Vinasse | 10 | 40 | 50 | 51.47 | 91.58 | |

Note: Abf II and Xyl III from H. insolens, Abf III from M. giganteus, β-xyl from T. reesei. The total amount of the water-soluble enzyme protein and water-insoluble substrate used is a 0.55 g/kg (DM), and the total amount of the enzyme used for the vinasse is 0.45∼0.55 g/kg (DM).
4.3.6. Enzyme Reactor
An enzyme reactor is the equipment used for the reactions between enzymes and substrates. Enzyme reactors can be divided into different types according to different methods. The reactor characteristics and their corresponding applications are listed in Table 4.2 (Lehe, 2006; Chen, 2008).
Table 4.2
Introduction of Enzyme Reactors
| Type | Operation mode | The main characteristics | |
| Homogeneous enzyme reactor | Agitator tank | Batch, batch | Using blender mixing |
| Ultrafiltration membrane reactor | Batch, batch and continuous | Membrane allows only low molecular compounds and enzyme pass through, suitable for large molecules substrates | |
| Immobilized enzyme reactor | The drum | Partial, half batch and continuous | Immobilized enzyme suspended in the solution |
| Fixed bed/packed bed | Continuous | ||
| Fluidized bed | Batch and continuous | ||
| Membrane reactor | Continuous | ||
| Bubbling tower | Partial, half batch and continuous | Suitable for the reactions with gas |

In practical applications, enzyme reactors are chosen based on the enzyme, substrate and product characteristics, operating conditions, and operation requirements. Free enzyme-catalyzed reactions are usually carried out in the stirred reactor; for the low yield but expensive enzymes, enzyme recovery is very important. So, it is very suitable to use a membrane reactor; for the enzyme reactions gas involved, we can choose the bubble reactors. For the high temperature resistance enzymes, it is better to choose the jet reactor.
If the molecular weight of the product is small, it is better to choose the membrane reactor, because micromolecules can easily pass through the membrane and get separated with the macromolecular substrate. However, the traditional membrane reactor has an obvious disadvantage: the concentration of the reducing sugar obtained is very low, so it is not suitable for the sequential treatment. Chen designed an ultrafiltration membrane bioreactor (Fig. 4.3) consisting of six enzymitic vessels which were used to increase the reducing sugar concentration. The effects of enzyme loading, the number of enzymatic vessel, and the dilution rate on enzymatic hydrolysis of steam-exploded rice straw were studied. The results show that the best condition is: 20 FPU/g, four enzymatic vessels, and 0.75 h−1 (D). Compared with the traditional batch reaction, the conversion of the substrate increased significantly from 18∼21% to 39.5%. Using the ultrafiltration membrane bioreactor consisting of one enzymatic vessel, the production of reducing sugar increased from 0.25 to 0.4 g/g, and the final reducing sugar concentration in the product steam increased from 4.56 to 27.23 g/L.

Figure 4.3 1, Thermostatic water bath; 2∼5, enzymatic membrane tank; 6, peristaltic pump 7, membrane module; 8, sugar tank; 9, pressure control valve.
4.4. Economic Analysis of Cellulase Production
4.4.1. Relations Between Particularity of Cellulose and Production Costs
As macromolecular proteins with catalytic activity, the function of cellulase and the stability of the integrity depends on its native conformation, which is very sensitive to temperature, the operating strength, and pH environment. Therefore, the properties put up high requirement on the cellulose production, which increases the production cost.
1. The low production from the producing strains
The current conversion rate of substrate of industrial strains is, on average, between 6% and 10%, which means that more than 90% of the raw material cannot be converted. Meanwhile, the enzymatic activity of native cellulose is low. The enzymatic activity of industrial grade is generally 2,000∼5,000 U/g. However, laboratory grade of cellulose activity is commonly about 10,000 U/g or more, indicating that there is plenty of scope for industrial fermentation to be improved.
2. Fermentation conditions
Different from the general chemical production, the cellulase fermentation requires high control of operation conditions, such as dissolved oxygen, temperature, pH, and other sophisticated requirements. Any unsuitable operation conditions will result in activity loss of the product. At the same time, the average growth phase of cellulase producing strain is 72 h and the average fermentation phase lasts about 4 d. The longer fermentation period leads to a continued consumption on sterile air, water, electricity, and manpower, which constitute an important part of production costs. In a production cycle, the fermentation process consumes most of the steam, water, electricity, and other energies.
3. Enzyme recovery
The enzyme recovery efficiency is also a key factor to determine the overall production cost. Currently, the enzyme recovery in industry production was 70% and the deactivation rate is about 10–30%. In other words, due to the limitations of the recovery techniques nowadays, 30–40% of cellulase cannot be converted into a commodity. Therefore, improving the separation efficiency methods is key to improve production efficiency and reduce production costs.
4.4.2. Relationship Between Process Indicators and Production Costs
In this section, we will take a cellulose factory with annual output of 480 tons as an example, to analyze the relationship between process indicators and production costs. The aim is to determine the limiting factors leading to high production cost of cellulase.
4.4.2.1. Relationship Between Production Costs and the Fermentation Period
Fermentation period is directly related to the cost of production; it is not only related to the annual production capacity of the factory, but also is the most important factor determining the consumption of water, electricity, labor and other expenses. The current average period of cellulase fermentation is four days (including seed cultivation, fermentation processes, products recovery, etc.). If using short-fermentation period strain or new control technology to compress the fermentation period, the cost of inputs can be greatly reduced.
4.4.2.2. The Relationship Between Fermentation Yield and Cost
Fermentation yield is also a key factor in the decision of cellulose production cost. The current conversation rate of substrate is about 6.5%. If this rate was increase 1–7.5%, raw material input costs can be greatly reduced. Table 4.3 shows the comparison of the raw material costing.
Table 4.3
List of Fermentation Yield and Cost Relationships
| Project | Name | The unit price | Consumption/t | Consumption/t | A total of 480 tons |
| The raw material | The raw material | 500 | 5.17 | 4.4807 | 165,432 |
| Wheat bran | 1,000 | 1.293 | 1.1206 | 82,752 | |
| Yeast extract | 1,500 | 0.012 | 0.0104 | 1,152 | |
| Lignocellulose | 2,800 | 0.24 | 0.208 | 43,008 | |
| Peptone | 2,500 | 0.072 | 0.0624 | 11,520 | |
| MgSO4 · 7H2O | 5,000 | 0.0072 | 0.0062 | 2,400 | |
| KH2PO4 | 8,000 | 0.096 | 0.0832 | 49,440 | |
| CaCl2 | 1,250 | 0.0072 | 0.00624 | 960 | |
| Tween-80 | 15,000 | 0.176 | 0.152533 | 169,440 | |
| Ammonium sulfate | 1,200 | 1.573 | 1.363267 | 120,960 | |
| Bubble enemy | 700 | 6.667 | 5.778067 | 299,040 |

If the conversion rate increased by 1%, as we can see, a cost of raw material of at least 2 million Yuan can be saved. At the same time, due to the reduction in the consumption of raw materials, we can save at least about 5,000 tons of water, 1.78 × 106 kw/h factory power, 7.4 × 104 ton steam, which add up to about 2.95 million Yuan in total and equal 19.6 and 10.2% of profit and total cost, respectively.
4.4.2.3. The Relationship Between Enzyme Activity and Production Costs
Compared with the conversion rate of the substrate, enzyme activity impacts great the production cost. If the enzyme activity were increased from the current average level (4600∼4700 U/g) by 1000 U/g, the total cost could be reduced by 3,000,000 Yuan (calculation method as aforementioned). In addition, the market price of cellulose shows a linear relationship with enzyme activity. In other words, if the current level of production (4600 U/g) increased to 5600 U/g, the market price will be higher than the original price 5600/4600 = 1.22 times, so profit growth will increase more than 1.22 times.
4.4.2.4. The Relationship Between the Filtration Method and the Production Cost
Product separation and purification is one of the major parts of the cost. For existing separation methods, the costs mainly arise from two aspects: raw materials and energy consumption.
First, in the enzyme recovery process, it requires ammonium sulfate to precipitate the enzyme cellulase. The total enzyme loss rate in this process is about 20% of the total enzyme. To product 480 t cellulose enzyme consumes at least 591 t ammonium sulfate, which is about 7.8% of the total cost. Feed cellulases must use ethanol as the precipitating agent. To product 480 t cellulose enzyme consumes at least 1870 t ethanol, which is about 21.4% of the total cost. Therefore, if we found a new separation method using a cheaper alternative to ammonium sulfate or ethanol, input costs could be greatly decreased.
Moreover, because the cellulase is very sensitive to the environment, if the separation conditions (temperature, pH, etc.) are not suitable, it will result in the loss of enzymatic activity. Under existing conditions, the enzyme recovery rate in the precipitation section is only 90%, the recovery rate of the filtering portion is 95% (the highest level), and the loss of the enzyme activity was 7–10%. Therefore, after the whole separation process, the loss rate of the enzyme is as high as 20–23%. If we can reduce the loss of the enzyme activity or increase recovery efficiency, the inputs will be greatly saved.
4.4.3. The Major Policy to Reduce Cellulose (or Conversion) Enzyme Economic Costs
Through the above analysis, it can be seen that the product cost can be saved from at least four aspects: (1) increase the rate of substrate conversion; (2) enhance the enzyme activity; (3) improve the product recovery; and (4) compress the fermentation cycle. Table 4.4 summarizes the achievements in enzyme development in recent years.
Table 4.4
The Main Strategy to Reduce the Cellulase Cost and the Corresponding Effects
| Source of enzyme gene | Expression host | Enzyme type | Enzyme activity |
| Thermonospora YX | E. coli | Endoglucanase | 5.8 μmol/min/mg × 103 |
| B. subtilisDR | E. coli | Endoglucanae | 0.82 U/mL |
| Xylella fastidiosa | E. coli | Endoglucanase | 2.39 μKat |
| Azoarcussp. strain BH72 | E. coli | Exoglucanase | 30 U/mg of protein |
| Ruminococcus flavefaciens | E. coli | Endoglucanase | 19.4 μg/min/mg |
| Bacillus sp. strain KSM-64 | B. subtilis | Endoglucanase | 21,700 U/L |
| Thermomonospora fusca | S. lividans | Endoglucanase | 10 U/mL |
| A. tubingensis | K. lactis | Endoglucanase | — |
| Cryptococcus sp. S-2 | P. pastoris | Endoglucanase | 4.36 U/mg of protein |
| T. reesei | P. pastoris | Exoglucanase-7 | — |
| T. reesei | P. pastoris | Exoglucanase-1 | — |
| T. reesei QM9414 | S. cerevisiae | Endoglucanase | — |
| T. reesei QM9414 | S. pombe | Endoglucanase | — |
| T. reesei | S. pombe | Endoglucanase-2 | Exoglucanase-3.81 U/mg of protein |
| T. reesei | Y. lipolytica | Endoglucanase-1 | Endoglucanase-0.1 U/mg of protein |
| A. fumigatus Z5 | P. pastoris | β-Glucosidase | 101.775.2 U/mg of protein |
| Candida wickerhamii | S. cerevisiae | β-Gluocosidase | — |
| P. chrysosporium | P. pastoris | β-Glucosidase | 52 U/mg of protein |
| Commercial mixture | Supplier | Reported enzyme activities (U/mL unless otherwise specified |
| Celluclast 1.5 L FG | Novozyme | 65 FPU, 12 β-glucosidase, 660 xylanase (bwx) 60.7 FPU, 6.5 β-glucosidase |
| Novozyme 188 | Novozyme | 8.5 FPU, 665 β-glucosidase, 123 xylanase (osx), 29.3 α-arabinofuranosidase, 16.6 β-xylosidase, 116 α-galactosidase, 0.6 feruroyl esterase (U/mg) 1 endoglucanase, 0.35 exoglucanase, 14.75β-glucosidase, 10 β-xylanase, 0.22 β-xylosidase, 0.09 α-arabinofuranosidase 0.1 FPU, 661 β-glucosidase |
| Spezyme CP | Genencor | 58.2 FPU/ml, 128 β-glucosidase, 2622 xylanase(osx), 22.6 α-arabinofuranosidase, 7.3 β-xylosidase, 0.39 α-galactosidase (U/mg) 1.4 FPU, 21.8 CMCase, 0.09 exoglucanase, 1.82 β-glucosidase, 15 β-xylanase, 0.56 β-xylosidase, 0.38 α-arabinofuranosidase, 55.2 FPU, 15.4 β-glucosidase |
| Multifect (xylanase) | Genencor | 0.77 FPU, 35.9 β-glucosidase, 25,203 xylanase (osx), 9.44 α-arabinofuranosidase, 22.6 β-xylosidase, 2.39 α-galactosidase, 1.3p-coumaroyl esterase (U/mg) 6.3 endoglucanase, 0.46 exoglucanase, 3.3 β-glucosidase, 209 β-xylanase, 4.9 β-xylosidase, 3.21 α-arabinofuranosidase |
| Accelerase 1000 | Genencor | 93 FPU, 7.3 CMCase, 1632 β-glucosidase, 849-xylanase 67.3 FPU, 84.2β-glucosidase |
| Primafast Luna CL | Genencor | Endoglucanases, no exoglucanase |
| GC220 | Genencor | 92.8 FPU, 99.7 β-glucosidase, 2782 xylanase 9(osx), 3.06 α-arabinofuranosidase, 7.3 β-xylosidase, 3.9 α-galactosidase |
| Shearzyme | Novozyme | 27 FPU, 5.0 β-glucosidase, 2293 xylanase (bwx) |
| NS50013 | Novozyme | 63 FPU, 8 β-glucosidase, 1117 xylanase (bwx) |


..................Content has been hidden....................
You can't read the all page of ebook, please click here login for view all page.