
Ecosystems and Biomass Systems
Hiroto Toda, Tomoe Shimizu, Jun Shimada and Takashi Motobayashi
Abstract
In this chapter, ecological and biomass systems of Satoyama, uplands, and paddy fields are introduced. Forests in the Satoyama region provide wood for fire logs to support life of nearby villagers and fallen leaves for compost to make surrounding farmland fertile. The Satoyama management practices also played an important role in maintaining the biodiversity and other important functions of the forests in the Satoyama region. In the section on long-term experimental upland fields, we discuss the impact of tillage and fertilizer application on the soil fauna through the soil detritus food web, and the role of soil fauna in crop production will be discussed. The Japanese paddy fields have provided food and a habitat for various organisms as well. However, the numbers of non-cultivated and/or abandoned paddy fields have increased recently. The discussion will also cover the possibility of utilizing these unused paddy fields to grow highly productive rice cultivars to supply a large amount of biomass and provide a habitat for various organisms.
Keywords
Biodiversity; biomass production; crop protection; food web; high-yielding rice; Satoyama system; self-fertilizing; soil fauna; soil management; symbiotic microbes
Chapter Outline
3.1. Natural Ecosystems and Satoyama Utilization
3.1.3. Sustainable Use and Management of Satoyama
3.2. Biomass Production in Cultivated Fields and Conservation of Ecosystems
3.2.1. Continuous Use and Ecosystems of Cultivated Fields
3.2.2. Detritus Food Web in Upland Fields
3.2.3. Effects of Manure Application on Biota in Upland Fields
3.3. Biomass Production and Biodiversity in Paddy Field Ecosystems
3.3.1. Rice Production in Paddy Fields Characteristic of Paddy Field Ecosystems
3.3.2. Biodiversity of Paddy Fields in Japan
3.3.3. Coexistence of Biomass Production and Maintenance of Biodiversity in Paddy Fields
3.1 Natural Ecosystems and Satoyama Utilization
Hiroto Toda
3.1.1 Forest Ecosystems
a Forest Productivity and Self-Fertilization
Organic materials such as fallen leaves and branches are continuously circulated in forests to add nutrients to the soil that will be taken up by growing plants. This is called self-fertilization of forests and maintains the productivity and soil fertility of forest land (Figure 3.1). Natural and artificial forests in deep mountains maintain nutrient cycling, or self-fertilization, unless they suffer huge natural disasters such as volcanic eruptions and landslides caused by earthquakes and/or heavy rain.
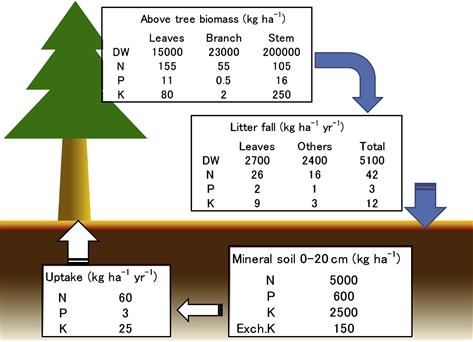
FIGURE 3.1 Nutrient circulation of Sugi (Japanese cedar; Cryptomeria japonica) and Hinoki (Japanese cypress; Chamaecyparis obtusa) artificial forests. Tokyo University of Agriculture and Technology, Field Science Center, Field Museum Ohya-san, the east of Gunma Pref., Japan. Data from Toda et al. (1991).
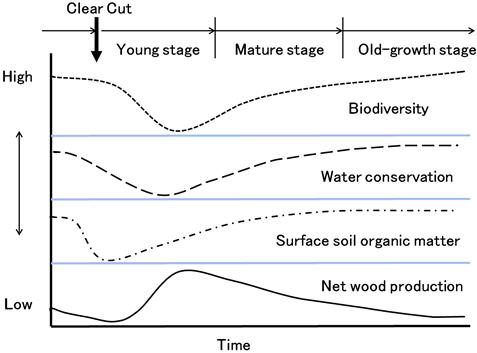
Cutting down trees for timber production disturbs nutrient cycling in artificial forests in addition to jeopardizing the functions of water conservation and biological diversity. Additionally, the tree cutting speeds up the decomposition of surface organic matter whereby a portion of carbon accumulated in forest soil is released into the atmosphere. However, these influences may be short-lived if the biological diversity as well as nutrient cycling and other public functions are restored to their former state by conducting proper forest regeneration and scheduling the cutting cycle at sufficient intervals (Figure 3.2).
b Forest Biomass Utilization in Deep Mountains
In Japan, the demand for timber increased rapidly from 1945 to 1965 because of the postwar reconstruction work after World War II. This paralleled the “fuel revolution” to replace fuel wood and charcoal with gas and petroleum. Hence, expansive forestation to plant Sugi (Japanese cedar; Cryptomeria japonica) and Hinoki (Japanese cypress; Chamaecyparis obtusa) artificially spread to deep mountains where deciduous broad-leaved trees had been cut down to make charcoal. As a result, Japan has 10 million hectares of Sugi and Hinoki forest resources in deep mountains for timber production.
Deep mountains have steep slopes with no modern forest road networks; high costs to transport wood from these deep mountains make logging unprofitable. However, logging for timber production along with utilizing the waste materials such as the slash produced during the sawing process is cost-effective and beneficial. However, the established Sugi and Hinoki forests accumulate a great amount of nitrogen and phosphorus with quantities equivalent to 4–7 years of the annual uptake. Hence, the leaves and branches after timber harvest should be left on the forest ground in order to maintain soil productivity (Toda, 2004).
c Huge Disturbance and Fertilizer Trees
When the disappearance of a large amount of forest vegetation occurs due to huge disturbances caused by natural disasters such as volcano eruptions, the restoration of forest ecosystems and ecosystem services is the first priority. On Miyake island, one of the Izu islands, forest ecosystem disturbances caused by Mt. Oyama’s eruptions have been replicated at an interval of about 20 years for many decades.
Ohbayashabusi (Alnus sieboldiana) is a popular pioneer tree in the Izu islands. It has numerous root nodules consisting of symbiotic bacteria with the capability of fixing nitrogen from the atmosphere. This species is capable of growing well in soil with poor nutrients (Toda et al., 2007, 2008). Ohbayashabusi originally grew in the Izu islands; the characteristics of the tree have been developed to replicate the forest in regions that have undergone a massive disturbance (Kamijo et al., 2002).
Ohbayashabusi is often used as a fertilizer tree for forests in Honshu because its leaves have a rich nitrogen concentration so that the leaf litter is capable of rapidly replenishing the soil nitrogen. The growth of this fertilizer tree declines after about 20 years, and the area is then replaced by subsequent stages of tree species. Therefore, fertilizer trees are useful to restore both forest vegetation and soil fertility. If fertilizer trees are properly managed, sustainable forest biomass resources and soil fertility can be maintained.
3.1.2 Satoyama Systems
a What is Satoyama?
Satoyama is a region with secondary forest and artificial forest surrounding a village. Compared with natural deep mountains, it is the secondary natural environment created and maintained by various human activities. Forests in Satoyama regions have long been the source of biomass for raw materials and energy such as fuel wood and charcoal; the fallen leaves are made into compost as fertilizer.
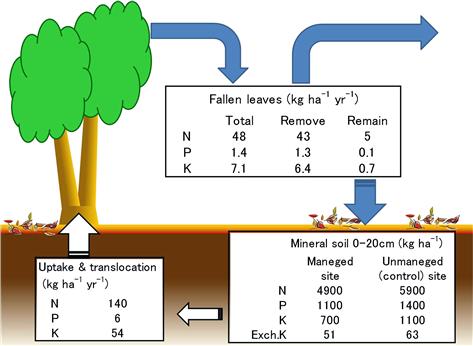
In the forests of Satoyama, harvesting wood and coppicing are done at a short rotation of 15–20 years because small trees with diameters of 5–20 cm are used to make fuel wood and charcoal (Figure 3.3). Also, the fallen leaves are collected and used as fertilizer for farmland so that the forest’s self-fertilization system is exploited. After decades of such management, the fertility of soil in Satoyama has been depleted (Figure 3.4). Therefore, trees that are capable of growing in soil are planted so that the forests in Satoyama now have strong regeneration power.
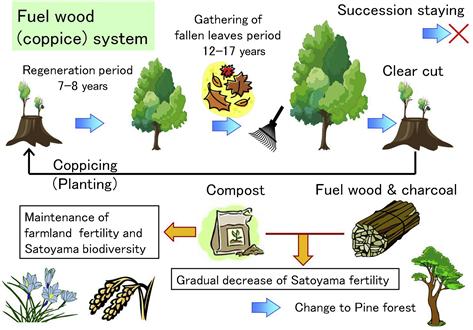
Trees growing in the secondary natural environment of Japan’s Satoyama include deciduous broad-leaved oak, i.e. Konara (Quercus serrata) and Kunugi (Quercus acutissima), and confiers, i.e. Akamatu (red pine: Pinus densiflora). When the practice of weeding and gathering fallen leaves is carried out regularly, the forest floor is incapable of supporting the growth of various plants inhabited by small animals. With the primeval natural forest in the deep mountains left untouched, Satoyama that was created by human activities has played an important role in maintaining the biological diversity of Japan.
b Strategy of Nutrient Acquisition in Satoyama Forests
Removing fallen leaves and cutting trees for short rotation periods have made Konara and Kunugi the major species growing in Satoyama because the short-term practice prevents the progressive growth of succession species. Repeated excessive cutting will degenerate the succession tree species; hence, Akamatu will become the predominant species (Figure 3.3).
Konara, Kunugi, and Akamatu can grow on such infertile land because ectomycorrhiza, which is a type of mycorrhizal fungi and has hypha tissue called Hartig nets (Hiroki, 2002) (Figure 3.5), have a symbiotic relationship with tree roots. Mycorrhiza in the host’s roots receives photosynthetic products generated by the host to grow, and supplies the host plant with nutrients and water gathered by its hyphae that extend under the ground (Box 3.1). The hypha is long and thin and has a larger surface area than the fine root of the host tree. In particular, mycorrhiza contributes much in promoting the uptake of phosphorus and building up tolerance of the host tree to resist drought and heavy metals (Smith and Read, 1997).

c Satoyama and Farmland Fertility
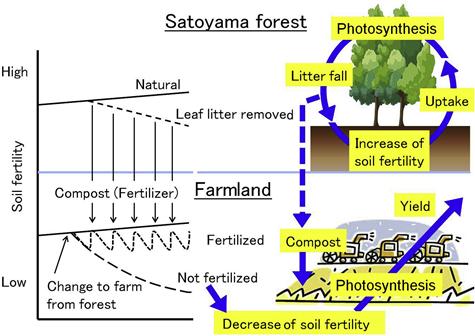
Gathering fallen leaves from Satoyama for providing nutrients to farmland has maintained the productivity of the farmland (Inui, 1996) (Figure 3.4). The ash after burning fuel wood and charcoal was also applied to farmland as soil conditioner or fertilizer.
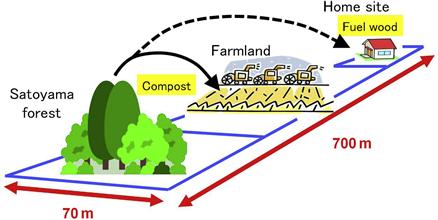
The San-tomi area located to the west of Saitama prefecture was reclaimed as an agricultural field in the Edo era. The land use pattern for home site, farmland, and Satoyama forest that adjoin one another on the rectangular 70 m × 700 m land (Figure 3.6) remains unchanged. The fallen leaves have been gathered and utilized to make compost on germination beds since around 1800, when sweet potatoes were extensively cultivated there. Results of the authors’ investigation reveal that the forest is a typical coppice of Musashino with Konara and a few Kunugi. About 10–30% of the annual nutrient uptake is estimated to be reduced by the gathering of fallen leaves in this managed Satoyama site (Figure 3.7). This site shows an obvious lower amount of soil nutrients than an adjoining control site where fallen leaves have not been gathered for several decades. However, a decline of Konara is not observed, although the gathering of fallen leaves has continued in the Satoyama forest. Hence, maintaining the growth of Konara should be suggested to enrich the soil with nutrients generated by mycorrhiza.
3.1.3 Sustainable Use and Management of Satoyama
a Crisis of Satoyama
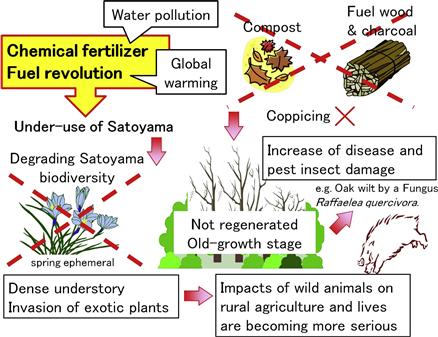
As mentioned before, intensive use of Satoyama with enhanced growth of fungi and production of edible wild plants helps the survival of spring ephemerals that has played an important role in restoring the original food culture and biodiversity in the Satoyama village. However, the use of Satoyama, such as leaf litter compost, fuel wood and charcoal, was decreased by the development of chemical fertilizer and the fuel revolution (Duraiappah et al., 2012) (Figure 3.8).
Satoyama forest became underused and the understory vegetation was altered by the advances of invading exotic bamboo species, as well as dense thickets and climbing plants. Because this alteration leads to changes in the forest floor environment from light-filled to dark space, the growth of spring ephemerals has declined (Figure 3.9). Wild animals of various sizes including deer, boars, and monkeys are also adversely affected due to the poor visibility in Satoyama. Human lifestyle is also affected because the buffer zone between wild animal habitats and human living space as well as many other functions of Satoyama have vanished. The unmanaged and ungenerating Satoyama forests have changed to a region with old growth of trees raged by diseases. Oak wilt, which is caused by a fungus, is spreading from Satoyama into deep mountain areas (Kuroda, 2001).

FIGURE 3.9 Comparison of understory managed site and unmanaged site in Motegi-mach’s Satoyama, east of Tochigi prefecture, Japan.
Wild animals, such as deer, boars and monkeys, enter the village due to the bad visibility in Satoyama.
Therefore, underuse of Satoyama forests has brought about many problems such as biodiversity degradation, a threat to rural lives caused by wild animals, spread of tree diseases, decrease of water conservation, and poor soil erosion control.
b Reconstruction of Satoyama Systems
The above observations illustrate the importance of protecting both the primeval nature and secondary natural environments such as the Satoyama area. As mentioned before, Satoyama biomass had been used and managed to provide compost and energy for a long period; it is used less nowadays because socio-economic conditions have become efficiency oriented. Most Satoyama forests have become unmanaged secondary forests, or have been replaced with artificial Sugi and Hinoki forests. The Satoyama Initiative to promote sustainable use and management of natural resources in human-influenced natural environments was implemented in 2010.
The woody materials of Satoyama forests are expected to be used as new economic resources for the development of biomass technologies in the coming years. For example, woody biomass has been used as pellets and other forms of fuel by local communities. In some areas, a system to recycle organic matter has been developed by compost manufacturing companies (Figure 3.10). Fallen leaves are gathered and packed in containers and shipped to recycling centers to be composted with other organic wastes, and then sold at a reasonable price to the farmers of the area.

FIGURE 3.10 Reconstruction of the Satoyama system with an organic recycling center in Motegi-mach, east of Tochigi prefecture, Japan.
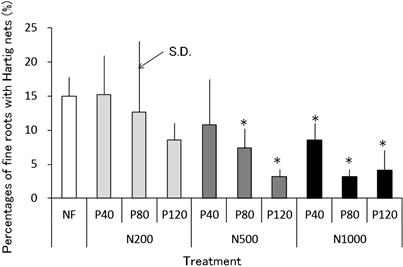
FIGURE 3.11 Percentages of fine roots with Hartig nets to total number of fine roots of Konara seedlings that were fertilized differently in soil.
NF: not fertilized (control) soil. N200, N500, and N1000: 200 mg N, 500 mg N, and 1000 mg N per 100 g fertilized soil respectively. P40, P80, and P120: 40 mg P, 80 mg P, and 120 mg P per 100 g fertilized soil respectively. ∗Significantly different from NF at P < 0.05 in t-test.
The capability of forest to absorb CO2 will be enhanced by practicing proper forest management, e.g. thinning Sugi and Hinoki forests and regenerating the Konara forest. These management practices produce woody biomass while contributing to the reduction of CO2 emissions. The national government and some regional governments have established systems for verifying the amount of CO2 absorption and emission reduction as credits, known as the “Offsetting Scheme (J-VER Scheme)”.
Reconstruction of the Satoyama system is a noteworthy model of local production for local consumption. Therefore, this practice should be greatly promoted to extend the experience on sustainable use and management of Satoyama to other forests.
3.2 Biomass Production in Cultivated Fields and Conservation of Ecosystems
3.2.1 Continuous Use and Ecosystems of Cultivated Fields
Cultivated fields in Japan are divided roughly into paddy fields and upland fields; both types of fields have efficient production of biomasses, including crops. Therefore, crops of the same line and the same variety are often grown in cultivated fields on a large scale by removing many other species that used to be components of the original ecosystem in order to protect the crop. In cultivated fields, because the intention is to keep the biota simple, and to keep succession at an early stage in cultivated fields, the cultivated field is essentially an unstable agricultural ecosystem. In addition, because the stem, leaf, root, and seed or fruit of crops is harvested and removed from the ecosystem, the material cycle is cut off. Hence, the current practice is to implement strong management based on recent advances in technology to maintain a stable ecosystem and sustainable cultivation of the field.
In order to utilize farmland sustainably, it is desirable to keep the soil environment the same before and after harvesting the crop. Compensating the loss of soil components absorbed by plants removed from the field as crops, and rapid decomposition of the organic matter (e.g. root secretions and crop residues) left in the field are essential. If these tasks are efficiently performed, “injury by continuous cropping” is alleviated. However, the technology to restore the soil conditions immediately after harvesting has not been developed at present. Hence, the problem of continuous cropping injury is avoided by the introduction of a crop rotation system by planting a different type of crop after some crops have been harvested. Monoculture systems in which only a single type of crop is planted may lead to outbreaks of particular insect pests or pathogens depending on the crop. The same is true for soil ecosystems; continuously cropping some plants causes the soil pest density to increase gradually and jeopardize the crop. As a result of injury by continuous cropping of a single type of crop, imbalance of soil nutrients is caused by the lack of certain nutrient components that have been depleted excessively but unnecessarily. The accumulation in cultivated soil of particular substances that inhibit plant growth, such as root secretions, have been considered. The injury caused by practicing continuous cropping does not happen in paddy rice fields even if the rice plant is grown every year because pathogenic microbes and soil nematodes cannot survive in the paddy soil that is submerged in irrigated water. A practice known as “paddy–upland rotation” can be implemented to use a cultivated field as a paddy rice field and an upland field alternatively. Because the species that can survive in the presence of irrigation water are different, paddy–upland rotation is effective for the control of weeds and pests.
Decreasing the energy input while maintaining or improving productivity is important to Japan’s agriculture. For cultivation of crops, much energy and advanced technology are needed to remove harmful components and maintain the condition of nutrient balance in the soil. The organism-preserving ability of the farming ground is a key to achieving these goals. However, because efficient production is pursued, the agricultural field is managed to exclude beneficial organisms for the sake of keeping only cultivated crop in the field without considering the benefit and damage to crops.
Soil is the site where various nutrients and water are supplied to support plant growth, and plant residues are resolved on-site to return nutrients to the field. In most ecosystems, the resources of the energy and nutrients originate from remains of flora and fauna that are resolved or decomposed by microorganisms, nematodes, annelids, and micro-arthropods living in the soil. Finally, the excrement and remains of these organisms are resolved by microorganisms to inorganic or organic nutrients that are available for plants to use. In the soil, there are numerous species of animals feeding on the fungi, bacteria, and the flora and fauna remains; these soil-dwelling animals play an important role in the resolution process in the ecosystem. Soil nematodes maintain the microbial population for a long time to speed up the resolution of an organism by microorganisms (Trofymow and Coleman, 1982); hence, there is a close relationship between the cluster structure of a nematode and soil fertility (Bongers, 1990).
The proliferation of microorganisms in soil is relatively slow. It is thought that small arthropods such as mites and collembolans activate microbes, expanding the surface area by fragmentation of litter to disperse microorganisms (Schowalter, 2000). Ants and earthworms mix the soil’s organic and inorganic components by feeding on the soil to increase the soil pore space (Crossley, 1977; Seastedt and Crossley, 1984; Whitford, 2000). Through those processes, soil animals and fauna have a significant direct and/or indirect impact on the capability of land to produce primary production.
In cropland, soil organisms play an important role in crop protection. Carabid beetles are not only the litter transformer in pedogenesis but also a generalist predator in the upland field. The generalist predators included in macrofauna suppress the density of pests in the crop field. The density of specialist predators and parasitoids that have high ability of finding and killing target pest is nearly dependent on the density of pest because of their feeding habit. Consequently, predators and parasitoids usually become populated in the cropland when the pest is abundant, and leave when the pest density is diminished. On the other hand, generalist predators can use various types of prey that survive in the vicinity of the field. Hence, they are regarded as the most important natural enemy to pests during the first period when the latter invade the field and propagate. Results of some research show that fungivorous nematodes, collembolans, and mites are effective in controlling plant diseases caused by fungi (Ishibashi and Choi, 1991; Nakamura et al., 1991; Matsuzaki and Itakura, 1992). Therefore, soil fauna may be an important factor for producing food safely and efficiently.
Since 1999, our research team started an investigation to clarify the role of soil fauna in upland ecosystems in Japan. The study is conducted in a long-term experimental upland field (1.5 ha area) established in 1992 for growing dent corn and soybeans alternately in summer and for growing grass in winter. It is located in Field Museum Fuchu, Field Science Center, Faculty of Agriculture, Tokyo University of Agriculture and Technology. The experimental field is divided into 20 plots to conform with the two-way ANOVA analysis of the results. Each plot is managed using different tillage methods, i.e. conventional tillage (T) or reduced tillage (RT), combined with various fertilizer applications, i.e. chemical fertilizer (F) or manure application (M). As a result of our research in the long-term experimental field, we will discuss the effects of soil management on the cropland food web based on our long-term field experimental results.
3.2.2 Detritus Food Web in Upland Fields
In terrestrial ecosystems, the main resources necessary for organisms to live in the soil are detritus, fragments of dead plant organic matter such as plant leaves, and roots’ peeling cells. Around the detritus, microorganisms and microphagous species form a complex degradation loop known as the “detritus cycle”, which is joined by higher predators or large soil animals that eat litter directly to form a soil detritus food. The detritus food web leads to the grazing food chain that plays an important role in supporting the food web of terrestrial ecosystems.
In the upland field ecosystem, any approach to till the soil for crop production disturbs the soil food web. Tillage will not only disrupt the habitat of soil organisms but also help microbial degradation by mixing the soil with crop residue. Introducing compost is a good way to invest organic resources into the detritus food web.
No tillage or reduced tillage management increases under-decomposed organic matter in the field and accumulates crop residues on the soil surface. Through the food web, tillage methods or manure application have a great impact on soil fauna.
In 1992, when the experimental field was first developed, there was no difference in the number of individual organisms in the tilled and untilled fields. In 1997 and 1999, reduced tillage plots had significantly higher populations of mites and collembola than fully tillage plots. The influence of fertilization is reflected by the number of individual organisms in 1999, when the populations of mites and collembola were significantly higher in manured plots than chemically fertilized plots. In 1999, research on the population of arthropods living on the ground and in crops was carried out. Although there is no statistical significance, numbers of both generalist predators (natural enemy) captured using pitfall traps and herbivores (insect pest) captured using Roth-type traps show some correlation with soil management. When the same tillage method was used, manured plots had larger populations of the predator and smaller populations of the herbivores than chemically fertilized plots. Also, an antagonistic relationship is observed between the populations of prey and predator (Figure 3.12).
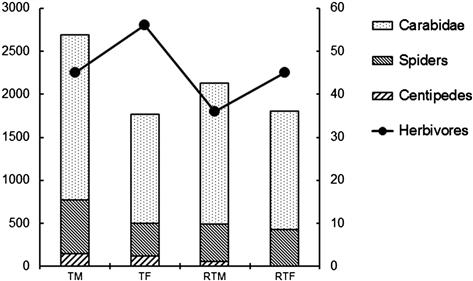
FIGURE 3.12 Number of individuals of prey and predator in each plot.
F: chemical fertilizer; M: manure; RT: reduced tillage; T: tillage. Bar graph presents the number of predators captured by pitfall trap and the solid line with filled circles the number of herbivores captured by Roth-type trap in the long-term experimental field during soybean cultivation period in 1999.
3.2.3 Effects of Manure Application on Biota in Upland Fields
a Microorganisms and Protozoa
Sato and Seto (2000) investigated the relationship between microbial biomass and method of tillage or fertilizer application. Their results show that continuous use of manure increases the fungi biomass although no difference is observed in the bacterial biomass. The order of fungi biomass is RTM > TM > RTF > TF. The continuous practice of manure application and reduced tillage is thus assumed to have a synergistic effect on biomass fungi.
Protozoa of heterotrophic and autotrophic species account for 70% of the total soil respiration rate by the soil fauna, 14–66% of the carbon mineralization, and 20–40% of the nitrogen mineralization (Griffiths, 1994; Ekelund et al., 2001). This fact allows us to assume that protozoa play an important role in crop production in upland ecosystems. A single ciliate can consume 300–400 individual bacteria whereas an amoeba can consume 2000–4000 bacteria (Ekelund and Ronn, 1994).
Griffiths (1994) reported that protozoa use approximately 40% of the nutrients absorbed from prey microorganisms for cell regeneration, and discharge the remaining 60% either directly or indirectly through respiration as inorganic components. The inorganic components released by protozoa will be used by microorganisms, plants, and animals. Concerning the role of protozoa in the rhizosphere, Clarholm (1994) considered that bacteria fixed on the surface of soil organic matter are in a steady state because they do not grow due to limited carbon source. Polysaccharide emitted from the tip of the roots and detached root cell during the growth of plant roots is easily biodegradable, and is used by the bacteria as a carbon source.
In addition, bacteria mineralize and uptake soil organic matter into cells as bacterial nitrogen; protozoa that are attracted to the CO2 released by bacteria feed on the bacteria to excrete inorganic nitrogen. Hence, bacteria-derived inorganic nitrogen becomes plant fertilizer near the roots, and in the presence of protozoa plant roots can intake the nitrogen efficiently due to the presence of bacteria.
Other organisms such as fungivorous nematodes, mites and collembolans, and protozoa contribute to the control of plant diseases because of their feeding habits. Foissner (1987) demonstrated that the inoculation of protozoa suppresses the potato soft-rot disease infected by Bacillus aroideae, and a genus of ciliate, Colpoda sp., inoculated to a cotton field decreases infection of wilt disease caused by Verticillium dahliae by 7–10% to increase the cotton yield by 30% (Foissner, 1987).
Wardle (1995) pointed out that the pressure exerted by protozoa on the bacterial biomass is higher than the pressure exerted by the fungivorous species on the fungus biomass in the detritus food chain. However, as described above, although the fungus biomass is significantly increased by the continuous use of manure, results of the long-term upland field study (Sato and Seto, 2000) do not show increasing bacterial biomass. We have investigated the number of individual protozoa living on these microorganisms in order to re-examine the effect of the difference in crop management on the number of bacteria and actinomycetes. The colony-forming units (CFUs) of aerobic bacteria and actinomycetes contained in 1 g dried soil were counted using a dilution plate technique using albumin agar medium. The protozoan population was enumerated using the most probable number (MPN) method designed by Ronn et al. (1995).
The proliferation of several typical ciliate and flagellate species among the species isolated from the upland field soil subject to long-term studies using the dilution plate technique were purified and subcultured in the albumin agar to enumerate their MPNs. Figures 3.13 and 3.14 show the CFUs of bacteria and actinomycetes respectively during the periods before and during soybean cultivation. Both manure and chemical fertilizer were applied on June 15; the soil was tilled on the same day, and seeded on June 18. CFUs of bacteria remain the same in the manured and fertilized plots. This observation is consistent with the results reported by Sato and Seto (2000). The influence of fertilization on the CFUs of bacteria appeared 10 days later and disappeared after 20 days. The transition density of the flagellate is shown in Figure 3.15. Ciliate density is about one hundredth of the flagellate and the same trend remains. Flagellate density rises from around 10 days after manure application to reach its highest level at 20 days. Despite an increase in bacterial population observed in the plots with chemical fertilizer application, the flagellate density is observed not to increase but decrease.
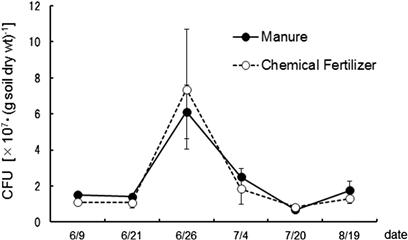
FIGURE 3.13 Number of colony-forming units of soil bacteria for albumin age medium during the soybean cultivation period in 2004.
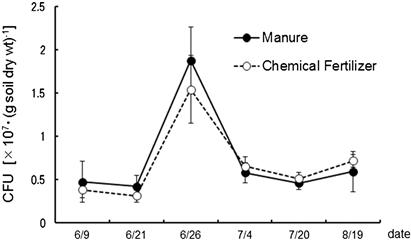
FIGURE 3.14 Number of colony-forming units of soil actinomycetes for albumin age medium during the soybean cultivation period in 2004.
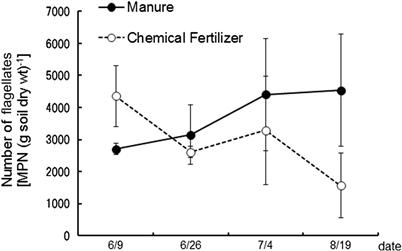
FIGURE 3.15 Number of soil flagellates counted using the MPN technique during the soybean cultivation period in 2004.
In general, introduction of the substrate through fertilization increases the initial population of microorganisms temporarily. However, because of some limiting factors such as substrate reduction and living environmental degradation, the microbial biomass has a tendency to decrease. Although there is a possibility that this observation only applies to our study, other factors such as weather conditions and tillage methods may also promote potential activities of bacteria and actinomycetes. Protozoa are sensitive to changes in the environment; they are transformed into a dormant cyst under unfavorable conditions. Typical culture methods cannot differentiate between cysts and active cells, whereas direct microscopy is incapable of observing small protozoa. To recognize the role of protozoa in upland fields, the development of molecular biology techniques and other ex-situ experimental model systems is needed.
b Soil Nematodes
In order to study soil nematodes, soil samples were collected from each plot, i.e. TM, TF, RTM, and RTF, of the long-term upland field where dent corn was planted. Two samples were collected at depths of 0–5 and 20–25 cm respectively from the surface. Nematodes were extracted from each sample using the Baermann funnel technique and classified into groups of free-living or plant parasites; their numbers were then counted. The nematode density was calculated and expressed as number of nematodes per gram of soil dried at 80°C for 48 h. The density of free-living nematodes in the soil layer at 20–25 cm depth is higher in the manure plot than the chemical fertilizer plot, and higher in the reduced tillage plot than the tillage plot (Table 3.1).
TABLE 3.1
Number of Nematode Individuals in Each Treatment Plot (mean ± standard deviation per gram of soil dry weight)
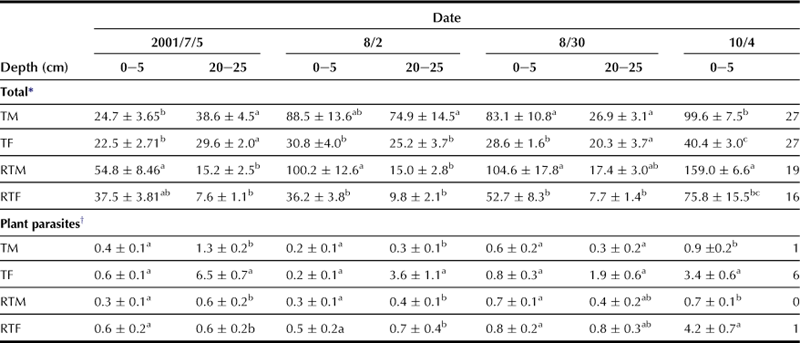
F: chemical fertilizer; M: manure; RT: reduced tillage; T: tillage.
There are significant differences between the different characters of the same sampling date (P < 0.05).
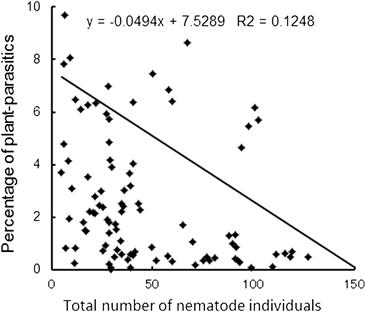
The observations demonstrate that manure application and reduced tillage management lead to a favorable soil environment for nematodes. On the contrary, the density of plant-feeding nematodes is higher in the chemical fertilizer plot than the manure plot, suggesting that the combination of conventional tillage and chemical fertilizer has a synergistic effect on the number of nematodes. Also, a significantly negative correlation exists between the density of total nematodes and the proportion of plant-feeding nematodes in each sample (Figure 3.16). These results suggest that an environment that has an increased number of free-living nematodes becomes unfavorable to plant-feeding nematodes.
c Soil Mesofauna
The effects of long-term manure application and reduced tillage in cultivated fields on the population dynamics of soil mesofauna were examined. Soil samples were collected from the four types of cultivated field, i.e. TM, RTM, TF and RTF, managed with different combinations of manure or chemical fertilizer application and conventional or reduced tillage. Soil mites and collembolans were extracted using Tullugren apparatus; they were classified into several groups mainly on the basis of family in order to enumerate the density for each group.
Fungivorous species of both mites and collembolans are dominant in every field. Densities of both mites and collembolans are higher in the manure plots than the chemical fertilizer plot. In particular, the plot with combined manure application and reduced tillage has remarkably higher densities of fungivorous mites and collembolans than plots with other combinations of nutrient application and tillage.
These results correlate positively with the fungal biomass in each treatment plot, and the density of predatory mites also correlates positively with the density of fungivorous species. Hence, manure application or reduced tillage will maintain the population of predatory macrofauna at a higher trophic level by increasing mites and collembolans that serve as the food for macrofauna.
d Soil Macrofauna and Insect Pests
Soil macro-arthropods and flying invasive insects were caught by using the pitfall trap or funnel-shaped trap in the above four types of field for enumeration, in addition to carrying out the aforementioned examination of soil mesofauna. The trapped organisms were classified into several functional groups based on their food habits before their numbers were enumerated.
The total number of arthropods tends to be greater in the field with reduced tillage than the conventionally tilled field, and greater in the field with manure application than the field with chemical fertilizer application. This tendency depends on the number of crickets (Gryllidae) or “other insects” that are neither herbivores nor predators. The number of phytophagous arthropods of two orders, i.e. Lepidoptera and Hemiptera, in the field with chemical fertilizer application is higher than in the field with manure application. Additionally, spiders, which have predatory food habits, are more numerous in the manured field than in the field with chemical fertilizer application. Few centipedes and earwigs (Labidura riparia japonica) were caught in the reduced tillage field, probably because of physical differences in the soil.
The number of phytophagous insect pests is reduced as a result of long-term manure application and reduced tillage. However, there is no correlation between the number of herbivores and that of predators. In nature, organisms may excrete volatile substances that function as allelochemicals to fend off natural enemies. In several cases, herbivorous insects were observed to perceive and avoid the volatile substances released from the general predaceous insect. Hence, such a relationship between organisms and their natural enemy, e.g. insect pests, may prevent the outbreak of specific insect pests that are harmful to crops. The manure application and the reduced tillage management are effective in maintaining the population of arthropods, which are prey of the general predaceous insects, so that sufficient allelochemicals will be produced by arthropods to prevent the density of harmful insects from becoming too high in the field.
3.3 Biomass Production and Biodiversity in Paddy Field Ecosystems
Takashi Motobayashi
Since paddy fields are constantly flooded with irrigation water, many characteristics (soil environment, biota, nutrient cycle, etc.) associated with paddy field ecosystems are different from those associated with upland field ecosystems. Paddy fields are typically puddled; the flooded soil is plowed and harrowed to destroy soil aggregates. Puddling eliminates water transmission pores, reduces the percolation rate, and drastically reduces gaseous exchange between the soil and the atmosphere. The puddling also results in the formation of a soil hardpan that restricts downward water flow to prevent the loss of nutrients by leaching. This practice also assists in transplanting rice seedlings and controlling weeds (Sharma and De Datta, 1985; Buresh and De Datta, 1990). Generally, rice plants are cultivated in paddy fields during late spring or early summer in Japan, and the fields are most likely irrigated with river water through channels during the growing season; during the fallow season, the irrigation water is drained from the paddy fields so that the paddy soil becomes dried. Therefore, the environment of wetland (such as shallow marsh) and upland appears periodically in paddy fields. The unique biological community established in the paddy fields has adapted to this rapidly changing environment. These communities in paddy field systems during rice plant growth and after rice harvest play an important role in building up and maintaining paddy field ecosystems.
3.3.1 Rice Production in Paddy Fields Characteristic of Paddy Field Ecosystems
a Nutrient Supply by Irrigation Water
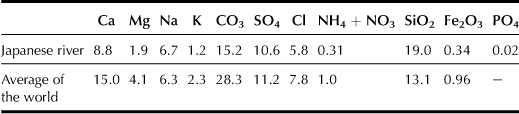
Japanese rivers, which tend to be shorter and swifter than continental rivers, contain relatively smaller amounts of various dissolved mineral nutrients as compared with the latter. However, silica is relatively abundant in Japanese rivers; many other types of dissolved mineral nutrients for rice plant growth are moderate (see Tables 3.2 and 3.3). These mineral nutrients accumulate in paddy soil and are absorbed by rice plants.
b Nutrient Supply and Cycling by Microorganisms (Photoplankton, Bacteria)
Soil fertility mainly depends on the availability of nutrient elements such as nitrogen, phosphate, and potassium contained in the soil. Among the major elements, nitrogen is of primary significance to affect crop production. The rice plant obtains a great portion of its nitrogen requirement from the organic nitrogen pool in the soil (Broadbent, 1979).
Nitrogen fixation is an important process to supply nitrogen for rice plants. Cyanobacteria and other bacteria (Azospirillum, Azotobacter, Clostridium, Beijerinckia, etc.) present in the paddy water or the paddy soil are capable of fixing nitrogen from the air contained in the paddy water (Firoza et al., 2002; Choudhury and Kennedy, 2004). The total nitrogen fixation capability of paddy fields has been estimated to be 1–6.8 g m−2 during the cropping season without artificial nutrient application (Hirano, 1958; Nishigaki and Shioiri, 1959).
Photoplankton takes in nitrogen and phosphate in paddy water and produces various organic substances. Although there is no exact information on the synthesis of photoplankton biomass in experimental plots, approximately 5–6 kg DW ha−1 is estimated to exist in the paddy field during the early stage of rice cultivation (Ichimura, 1954; Kimura et al., 2004).
Actually most organic matter applied artificially to the paddy soil annually is residue of weeds and rice stubble. The amount of weed and rice stubble biomass that was incorporated by spring plowing ranges from 1300 to 2300 kg DW ha−1 and 930 to 1500 kgDW ha−1 respectively (Kimura et al., 2004).
As mentioned above, many types of nutrients for rice plants are stored in the paddy soil as organic matter; it is then decomposed by various microorganisms, mainly bacteria, in the paddy soil. Because of a shortage of oxygen in the paddy soil caused by flooding water, the decomposition rate of the organic matter in the paddy soil during the flooding period is slower than that in the upland fields. As a result of accumulation of organic matter in the paddy soil, the surplus nutrients are recycled back to the paddy water while maintaining the fertility of the paddy soil so that outflow of the surplus nutrients from the paddy field to rivers and lakes is prevented.
c Nitrogen Metabolism During Flooding Season
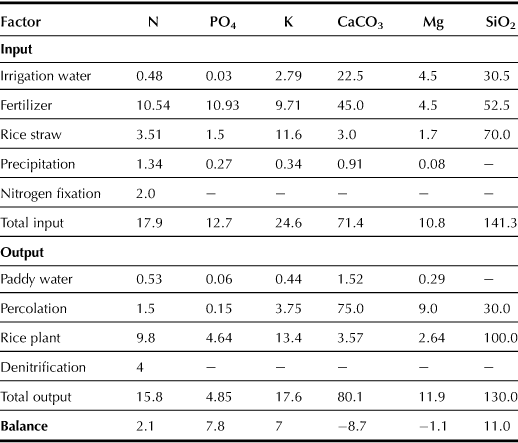
During most flooding periods, paddy fields develop a “reduced soil layer” as a result of active microbial metabolism. Surface paddy water contains oxygen dissolved from the atmosphere or produced by the photosynthetic activities of hydrophytes. This oxygen is supplied to surface soil through diffusion or with the downward movement of water; an “oxidized soil layer” thus develops in the uppermost part of the paddy soil (Takai and Kamura, 1966). The thickness is generally a few millimeters for the oxidized soil layer and 10–15 cm for the underlying reduced soil layer. The rice plant obtains a great part of its nitrogen requirement from the organic nitrogen pool of the soil (Broadbent, 1979). The organic nitrogen contained in the soil is mineralized to ammonium (NH4+) by microorganisms; the ammonium is stable under reduced soil conditions and adheres to the soil particles. Rice plants take up ammonium from the soil by absorption through their roots. However, if the ammonium exists in the oxidized soil layer, it is first converted to nitrite (NO2−), primarily by ammonia-oxidizing bacteria, and then oxidized into nitrate (NO2−) by nitrite-oxidizing bacteria. Nitrate is more soluble in water than nitrite; it moves from the oxidized soil layer to the reduced soil layer with the downward movement of the water. In the reduced soil layer, nitrate is denitrified by denitrifying bacteria into nitrogen gas (N2), which eventually escapes to the atmosphere in a process known as denitrification (Araragi, 1978). The rice paddy soil is known to have strong denitrifying activities (Nishimura et al., 2004); hence, about 20–25% of nutritional nitrogen for rice plant growth is lost to the atmosphere (Broadbent and Tusneem, 1971; Koyama et al., 1973) (see Table 3.3). However, application of nitrogen fertilizer to the reduced soil layer can suppress the microbial denitrification and improve the efficiency of fertilizer. Therefore, the uniform application of fertilizer to topsoil has been developed and implemented extensively, with the excess nitrogen being removed from the irrigation water by denitrification. Thus, some research on using paddy fields as a system to treat nutrient-polluted river water has been conducted (Zhou and Hosomi, 2008).
d Injury by Continuous Cropping
The injury caused by continuous cropping is a serious problem for upland field crops. It is caused by the deficiency of particular nutrients in the soil, accumulation of toxic substances in the soil, and prolific growth of soil-borne pest organisms (pathogenic microorganisms, nematodes) in the fields stimulated by continuous cropping. In particular, the increase of soil-borne pest organisms is the most severe problem caused by continuous cropping (Sekiya, 1992).
The soil in upland fields is continuously aerobic. Thus, aerobic microorganisms including plant pathogenic fungi or nematodes dominate in the community of microorganisms inhabiting the soil. As a result, crops cultivated continuously in the same field may be vulnerable to infestation by soil-borne pathogens.
As mentioned above, the reduced soil layer develops in the paddy soil during the flooding period, whereas the oxidized soil layer develops during the non-flooding period. Therefore, during the flooding period, aerobic microorganisms including plant pathogenic microorganisms become extinct or inactive, and anaerobic microorganisms dominate in the soil microorganism community in the paddy field. During the non-flooding period, anaerobic microorganisms are replaced by aerobic microorganisms in the soil microorganism community.
Thus, a particular soil-borne pathogenic microbial population may not develop continuously in paddy fields. Moreover, the accumulation of toxic substances in the soil may be suppressed by flooding water. Therefore, rice plants may be cultivated in the same paddy field continuously without causing injury by continuous cropping. The Japanese paddy field system is thus considered an effective and sustainable production system because nutrients are derived from irrigation water and converted into appropriate forms by various microorganisms. Therefore, the paddy field achieves effective recycling of surplus nutrients in paddy water and suppression of injury by continuous cropping. However, the paddy fields may operate more effectively if rotation cropping of rice and other field crops (such as soybean, wheat, vegetables) is implemented (Sekiya, 1992).
3.3.2 Biodiversity of Paddy Fields in Japan
For a long time in Japan, the paddy field has played important roles as agricultural land to produce rice and also to provide a habitat for various animals and plants. Example species and numbers of animals and plants detected in paddy fields are discussed in the following section.
Kobayashi et al. (1973) reported that 450 invertebrate species were detected in paddy fields in the Tokushima prefecture; 11 species of frogs were reported to use paddy fields as a breeding site (Maeda and Matsui, 1989). Additionally, 24 species of fish (Saito et al., 1988), 101 species of bird (Hidaka, 1998), and 186 species of plants (Kasahara, 1951) live in paddy fields either temporary or continuously. The paddy field ecosystem has more abundant species richness or species diversity than any other ecosystem. However, these data may have methodological or taxonomical problems (Hidaka, 1998).
Moriyama (1997) hypothesized that Japanese paddy fields were mostly developed in riverside wetlands; however, Japanese rice cultural systems had maintained the environment of the original wetlands before paddy fields were developed. Therefore, the paddy field has played a role of alternative habitats for organisms originally living in shallow wetlands located along the riverside.
Hidaka (1998) indicated that the Japanese paddy fields are temporal and spatial stable wetlands, and have higher sustainability than the original wetlands. He also suggested that there are great varieties in the time to flood the paddy fields, water content of paddy soils before flooding among various paddy fields, and various species may have been adapted to take advantage of these diverse paddy environments.
From the viewpoint of the life cycle of species using paddy fields, some species complete their life cycle in the paddy field; however, most species need the surrounding environments such as irrigation ponds and forests or groves for completing their life cycle. For example, Sympetrum, a dragonfly, uses paddy fields to lay eggs in autumn; the eggs hatch at puddling time next spring, and the larvae grow in paddy water during spring and early summer. Adult dragonflies then move from the paddy to adjacent or highland forests to live through summer. At the beginning of autumn, they return to paddy fields for breeding (Taguchi and Watanabe, 1985).
Hibi et al. (1998) and Saijo (2001) indicated that many species of predatory aquatic insects such as water stick, which is a Japanese water scavenger beetle, live in irrigation ponds adjacent to paddy fields. In other words, the adults of these insects migrate from ponds to puddled paddy fields for breeding and oviposition. The larvae grow in paddy water, where they prey upon abundant spices such as small invertebrates, tadpoles, and small fishes during summer. Adult dragonflies of the next generation return to irrigation ponds to hibernate at the beginning of autumn.
The landscape of “Satochi-Satoyama” consists of heterogeneous ecosystems including paddy fields, ridges between paddy fields, rivers, channels, forests, and upland fields. This complicated and diverse environment is one of the primary factors supporting various species in the paddy fields in Japan (Hidaka, 1998; Kiritani, 2000).
The total area of paddy fields in Japan is 2,600,000 ha, which accounts for 7% of the nation’s rural area and 20% of total flat land. Thus, the conservation of biodiversity in paddy fields is very important to conserve the biodiversity in Japan.
However, the serious problem caused by the disappearance of various species, especially aquatic species, inhabiting the paddy fields in Japan has been recognized recently. Causes of this disappearance include the drier paddy soils during the non-flooding period, disappearance of irrigation ponds due to adjustment of paddy fields, pollution of irrigation water by excessive use of pesticides and/or chemical fertilizers, acceleration of cultivation period of rice, and increasing non-cultivated or abandoned paddy fields. Hidaka (1998) has classified the rice cultivation practices in relation to the temporal and spatial stability of paddy fields. He suggested that the present paddy field environments are becoming unstable temporally and spatially, and therefore some permanent residents such as fishes and shellfishes that are vulnerable to unstable environment cannot survive in those unstable paddy field environments. Moreover, he hypothesized that migrants including Japanese crested ibis, stork, Japanese water beetle and giant water bug are more resistant to the unstable environment but they do not inhabit the paddy field. This is because their living cannot be supported by the decreasing population of permanent resident species in paddy fields. However, there has been little research conducted on the relation between the stability of paddy field environments and the population of individual species populations. More studies are needed on this in the future.
3.3.3 Coexistence of Biomass Production and Maintenance of Biodiversity in Paddy Fields
a Increasing Non-Cultivation and Abandoned Paddy Fields
Rice consumption has been declining in Japan, and rice production has been restricted to a balanced rice supply and demand since 1970. The rice production adjustment policy has been continued by the National Government. In 2011, the area of the paddy fields without rice cropping reached about 900,000 ha (MAFF, 2012a) that is about 35% of total paddy fields in Japan.
The declining and aging farmer population in recent years causes a substantial number of paddy fields, especially those located in mountainous and hilly areas, to be laid fallow (MAFF, 2010). On the other hand, diversion of agricultural lands to residential or business sites and the abandoned paddy fields are increasing in suburban areas (see Figures 3.17 and 3.18).


As mentioned earlier, the Japanese paddy field system is an effective and sustainable food and biomass production system, and increasing fallow paddy fields implies that a large portion of the agricultural production potential in Japan is lost.
Moreover, most of the non-cultivation or abandoned paddy fields are not periodically flooded and the aquatic organisms living in paddy fields disappear. Thus, increasing non-cultivation or abandoned paddy fields is becoming one of the serious problems causing declining biodiversity in Japanese paddy ecosystems to adversely affect the national land conservation (flood control, soil conservation) and maintenance of the agricultural landscape in Japan.
b Production of Whole Crop Silage (WCS), High-Yielding Rice in Non-Cultivation Rice Paddy
As mentioned in the above section, the area of paddy fields without rice cropping has reached about 1,000,000 ha. On the other hand, the percentage of self-sufficient food supply is only 39% on a calorie basis in Japan (MAFF, 2012b). Furthermore, 74% of the feed for livestock is imported from overseas (MAFF, 2012c). This striking shortage of self-sufficient supply against demand of feed causes serious problems for the livestock industry, similar to other problems including damage to the environment by animal waste and the risk of livestock diseases through imported feed. The enhancement of self-sufficient feed supply and the establishment of a recycling system between the livestock industry and agriculture are strongly recommended in order to solve these problems. The Ministry of Agriculture, Forestry and Fisheries (Japan) started a research project in 1999 to utilize rice as whole crop silage (WCS). As a result of this project, some new rice cultivars that have high productivity and are suitable for growing low-cost rice grain for WCS uses have been developed (Sakai et al., 2003). The area of paddy field cropping for WCS reached about 23,000 ha in 2011 (MAFF, 2012d). Furthermore, the production of rice grain for livestock feed has been increased. The area of paddy field cropping for this type of rice reached about 34,000 ha in 2011 (MAFF, 2012c).
Incidentally, bioethanol is widely used in Brazil and the USA as a biofuel additive for gasoline. The bioethanol can be made from very common crops such as corn and sugar cane. However, there have been concerns about producing ethanol fuel from food crops because the competition between bioethanol and food production may cause food shortages, especially for poor nations, in the future.
In Japan, because rice production has been in excess, using the surplus rice grain as raw material for bioethanol seems to be logical for the time being but may lead to future shortage problems. Hence, developing cellulosic ethanol production technique using rice straw for bioethanol production will be a preferred alternative method to produce bioethanol.
Thus, the rice cropping for WCS or bioethanol in non-cultivation or abandoned paddy fields may sustain the productive potential of Japan’s paddy fields in addition to conserving habitats for organisms inhabiting the paddy fields. Additionally, this practice may also be an effective means for maintaining the national land conservation and agricultural landscape in Japan.
c Subjects of WCS and High-Yielding Rice Production
At present, the production cost of WCS is 80 yen kg−1, twice as much as the cost of importing hay from overseas. The rice grain production cost is 133 yen kg−1, which is about three times the cost of importing corn from overseas. Thus, the breeding of new cultivars that have higher-yielding grain or whole biomass will be one of the primary technological innovations for achieving this objective (Horie, 2009). Nemoto (2010) indicated that, in addition to high yielding, the cultivars also need to be lodging resistant, disease resistant, insect and pest resistant, and suitable for direct sawing.
Moreover, with regard to rice cultivation practices for low-cost production, direct sawing that reduces the use of chemicals such as fertilizers and pesticides needs to be developed by conducting future research. The construction of a nutrient recycling system including paddy fields and livestock is also needed.
References
1. Araragi M. Denitrification in paddy field. In: Kawaguchi K, ed. Soil science of paddy field. Tokyo: Kobunnsha; 1978;256–263. [in Japanese].
2. Bongers T. Comparison of soil surface arthropod populations in conventional tillage and old field systems. Agro-Ecosystems. 1990;8:247–253.
3. Broadbent FE. Mineralization of organic nitrogen in paddy soils. In: Nitrogen and rice. Los Banos, Philippines: IRRI; 1979;105–118.
4. Broadbent FE, Tusneem ME. Losses of nitrogen from some flooded soils in tracer experiments. Soil Science Society of America Proceeding. 1971;35:922–926.
5. Buresh RJ, De Datta SK. Denitrification losses from puddled rice soils in the tropics. Biology and Fertility of Soils. 1990;9:1–13.
6. Choudhury ATMA, Kennedy IR. Prospects and potentials for systems of biological nitrogen fixation in sustainable rice production. Biol Fertil Soils. 2004;39:219–227.
7. Clarholm M. The microbial loop in the soil. In: Ritz K, Dighton J, Giller KE, eds. Beyond the biomass compositional and functional analysis of soil microbial communities. Chichester: John Wiley; 1994;221–230.
8. Crossley Jr DA. The roles of terrestrial saprophagous arthropods in forest soils. In: Mattoson WJ, ed. The role of arthropods in forest ecosystems. New York: Springer; 1977;49–56.
9. Duraiappah AK, Nakamura K, Takeuchi K, Watanabe M, Nishi M, eds. Satoyama-Satoumi ecosystems and human well-being: Socio-ecological production landscapes of Japan. Tokyo: United Nations University Press; 2012.
10. Ekelund F, Ronn R. Notes on protozoa in agricultural soil with emphasis on heterotrophic flagellates and naked amoebae and their ecology. FEMS Microbiology Reviews. 1994;15:321–353.
11. Ekelund F, Ronn R, Griffiths BS. Quantitative estimation of flagellate community structure and diversity in soil samples. Protist. 2001;152:301–314.
12. Firoza A, Zakaria ZA, Tahmida ZN, Mondal R. Increase in macro-nutrients of soil by the application of cyanobacteria in rice production. Pakistan Journal of Biological Sciences. 2002;5:19–24.
13. Foissner W. Soil protozoa: Fundamental problems, ecological significance, adaptations in ciliates and testaceans, bioindicators, and guide to the literature. Proger Protistology. 1987;2:69–212.
14. Griffiths BS. Soil nutrient flow. In: Darbyshire JF, ed. Soil protozoa. Wallingford, Oxon: CAB International; 1994;65–91.
15. Hibi N, Yamamoto T, Yuma M. Life histories of aquatic insects living in man-made water systems located around paddy fields. In: Ezaki T, Tanaka T, eds. The preservation in water environments: From the viewpoint of biodiversity. Tokyo: Asakura shoten; 1998;111–123. [in Japanese].
16. Hidaka K. The biodiversity and the preservation in paddy fields. In: Ezaki T, Tanaka T, eds. The preservation in water environments: From the viewpoint of biodiversity. Tokyo: Asakura shoten; 1998;125–148. [in Japanese].
17. Hirano T. Studies on blue green algae (part 2) Study on the formation of fumus due to the growth of blue green algae. Bulletin of the Shikoku Agricultural Experiment Station. 1958;4:63–74 [in Japanese with English summary].
18. Hiroki S, ed. Ecology of Satoyama. Nagoya: Nagoya University Press; 2002.
19. Horie T. Reductionistic and integrated approach in agricultural research. Japanese Journal of Crop Science. 2009;78:399–406.
20. Ichimura S. Ecological studies on the plankton in paddy fields I Seasonal fluctuations in the standing crop and productivity on plankton. Japanese Journal of Botany. 1954;14:269–274.
21. Inui T. The history and use of lowland forest in Kanto plain. Shinrin-Kagaku. 1996;18:15–20.
22. Ishibashi N, Choi DR. Biological control of soil pests by mixed application of entomopathogenic and fungivorous nematodes. Journal of Nematology. 1991;23:175–181.
23. Kamijo T, Kitayama K, Sugawara A, Urushimichi S, Aasai K. Primary succession of the warm-temperate broad-leaved forest on a volcanic Island, Miyake-jima, Japan ‘jointly worked’. Folia Geobotanica. 2002;37:71–91.
24. Kasahara Y. Studies on the number of arable weeds and their distribution in Japan Part IV. Nogaku- kenkyu. 1951;39:143–154 [in Japanese].
25. Kimura M, Murase J, Lu Y. Carbon cycling in rice field ecosystems in the context of input, decomposition and translocation of organic materials and the fates of their end products (CO2 and CH4). Soil Biology and Biochemistry. 2004;36:1399–1416.
26. Kiritani K. Integrated biodiversity management in paddy fields: Shift of paradigm from IPM to IBM. Integrated Pest Management Reviews. 2000;5:175–183.
27. Koyama T, Chammek C, Niamsrichand N. Nitrogen application technology for tropical rice as determined by field experiements using 15N tracer technique. Technical Bulletin (Tropical Agr Res Cen Tokyo). 1973;3.
28. Kobayashi T, Noguchi Y, Hiwada T, Kanayama K, Maruoka N. Studies on the arthropod associations in paddy fields, with particular reference to insecticidal effect on them I General composition of the arthropod fauna in paddy fields revealed by net-sweeping in Tokushima Prefecture. Kontyu. 1973;41:359–373 [in Japanese with English summary].
29. Kuroda K. Response of Quercus sapwood to infection with the pathogenic fungus of a new silt disease vectored by the ambrosia beetle Platypus quercivorus. Journal of Wood Science. 2001;47:425–429.
30. Maeda N, Matsui M. Frogs and toads of Japan. Tokyo: Bun-ichi Sogo shuppan; 1989; [in Japanese with English summary].
31. MAFF (2010). <http://www.maff.go.jp/e/annual_report/2010/pdf/e_all.pdf>.
32. MAFF (2012a). <http://www.e-stat.go.jp/SG1/estat/List.do?lid=000001087149>.
33. MAFF (2012b). <http://www.maff.go.jp/j/press/kanbo/anpo/pdf/120810-01.pdf>.
34. MAFF (2012c). <http://www.maff.go.jp/j/chikusan/sinko/lin/l_siryo/pdf/haigou_2409.pdf>.
35. MAFF (2012d). <http://www.maff.go.jp/j/chikusan/kikaku/lin/pdf/megru_1207.pdf>.
36. Matsuzaki I, Itakura J. Effect of collembola grazing on the reduction of rhizoctonia damping-off of vegetable seedlings in three types of toils under the laboratory conditions. Annual Report of the Society of Plant Protection of North Japan. 1992;43:133–134.
37. Moriyama H. What is protecting paddy fields? Tokyo: Nobunkyo; 1997; [in Japanese].
38. Nakamura Y, Itakura J, Matsuzaki I. Mycophagous meso soil animals from crop fields in Fukushima Pref. Edaphologia. 1991;45:49–54 [in Japanese].
39. Nemoto H. The status and subjects of rice production for bio-ethanol materials in Japan. Japanese Journal of Crop Science. 2010;79:224–226.
40. Nishigaki S, Shioiri M. Nitrogen cycles in the rice field soil: The effect of blue–green algae on the nitrogen fixation of atmospheric nitrogen in the water-logged rice soils. Soil Plant Food. 1959;5:36–39.
41. Nishimura S, Sawamoto T, Akiyama H, Sudo S, Yagi K. Mathane and nitrous oxide emissions from a paddy field with Japanese conventional water management and fertilizer application. Global Biological Cycles. 2004;18:GB2017.
42. Ronn R, Ekelund F, Christensen S. Optimizing soil extract and broth media for MPN-enumeration of naked amoebae and heterotrophic flagellates in soil. Pedobiologia. 1995;39:10–19.
43. Saijo H. Seasonal prevalence and migration of aquatic insects in paddies and an irrigation pond in Shimane Prefecture. Japanese Journal of Ecology. 2001;51:1–11 [in Japanese with English summary].
44. Saito K, Katano O, Koizumi K. Movement and spawning of several fresh water fishes in temporary waters around paddy fields. Japanese Journal of Ecology. 1988;38:35–47 [in Japanese with English summary].
45. Sakai M, Iida S, Maeda H, Sunohara Y, Nemoto H, Imbe T. New rice varieties for whole crop silage use in Japan. Breeding Science. 2003;53:271–275.
46. Sato A, Seto M. Fungal or bacterial biomass in volcanic soil with various land used and soil treatments (in Japanese). Soil Microorganisms. 2000;54:23–30.
47. Schowalter TD. Decomposition and pedgenesis. In: Insect ecology. New York: Academic Press; 2000;361–388.
48. Seastedt TR, Crossley Jr DA. The influence of arthropods on ecosystems. Bioscience. 1984;34:157–161.
49. Sekiya S. Function of paddy field. Tokyo: Ienohikari-kyoukai; 1992; [in Japanese].
50. Sharma PK, De Datta SK. Effect of puddling on soil physical properties and processes. In: Soil physics and rice. Los Banos: Philippines; 1985;217–234. IRRI.
51. Smith SE, Read DJ. Mycorrhizal symbiosis. San Diego: Academic Press; 1997.
52. Taguchi M, Watanabe M. Ecological studies of dragonflies in paddy fields surrounded by hills IV Spatial distribution of Sympetrum eroticum eroticum in relation to the seasonal fluctuations of shaded area. Bulletin of the Educational Faculty of Mie University. 1985;38:57–67 [in Japanese].
53. Takai Y, Kamura T. The mechanism of reduction in waterlogged paddy soil. Folia Microbiology. 1966;11:304–313.
54. Takayama S. “Guidebook of chronological science table” Natural history of river. 1986; Maruzen, Tokyo, pp. 160–161 (in Japanese).
55. Toda H. Effects of slash-removal on the nutrients circulation in forest ecosystems. Shinrin-Kagaku. 2004;40:33–38.
56. Toda H, Haibara K, Arai M. Nutrient circulation of a small watershed under an established Sugi (Cryptomeria japonica) and Hinoki (Chamaecyparis obtusa) stand. Bulletin of Experiment Forest, Tokyo University of Agriculture and Technology. 1991;28:1–22.
57. Toda H, Hanaoka K, Kishimoto K, Haibara K, Kametani Y. Mapping and mechanisms of acid buffer effect in volcanic ash on surface soil in Miyake Island. Journal of Japanese Society for Revegetation Technology. 2007;33:21–26.
58. Toda H, Hanaoka K, Ebara M, et al. Effects of forest soil properties on revegetation after volcano eruption 2000 in Miyake Island. J Jpn Soc Reveget Tech. 2008;34:21–26.
59. Trofymow JA, Coleman DC. The role of bacterivorous and fungivorous nematodes in cellulose and chitin decomposition. In: Freckman DW, ed. Nematodes in soil ecosystems. Austin, TX: University of Texas Press; 1982;117–138.
60. Wardle DA. Impacts of disturbance on detritus food webs in agro-ecosystems of contrasting tillage and weed management practices. In: Begon M, Fitter AH, eds. New York: Academic Press; 1995;105–185. Advances in ecological research. Vol. 26.
61. Whitford WG. Keystone arthropods as webmasters in desert ecosystems. In: Coleman DC, Hendrix P, eds. Invertebrates as webmasters in ecosystems. Wallingford, Oxon: CAB International; 2000;25–42.
62. Zhou S, Hosomi M. Nitrogen transformations and balance in a constructed wetland for nutrient-polluted river water treatment using forage rice in Japan. Ecological Engineering. 2008;32:147–155.