
Chapter 5
Primate Numbers
Introduction: Who’s on First?
In Chapter 2 we examined two energy forms that reveal much of our world to us, electromagnetic waves and mechanical waves, and then we discovered how eyes and ears evolved an ability to perceive this world. Along the way, we discovered that not only did our ears evolve to perceive mechanical waves in the form of sound, but that they had a peculiar ability to apprehend time and directly connected to the emotional centers of our brains. In Chapter 3 we examined that unique relationship of our ears to our perception of time, and then discovered that time itself was something of a moving target. Time is not absolute as we often believe, but relative to each individual. We proposed then, that by manipulating sound we would be able to manipulate both a person’s emotional state and their perception of time; in short, we would be able to manipulate their conscious experience and transport them into other worlds.
In Chapter 4 we described music as a tool we use to manipulate a person’s perception of time, and along the way, incite, arouse and immerse rather than communicate. We set out to define music more precisely when we use the term in this book. We now continue our journey through time, and begin to look for early signs of the emergence of music, language and theatre in mammals. In music, we will be looking for clues that will lead us to the creation of an experience that is artistic, temporal and non-referential, and probably involves both sound and sight. We are still many millions of years before such an experience can legitimately be identified as an attempt at art. Still, in the next couple of chapters, we will find surprising signs in the evolution of mammals, primates, and early humans that just such an evolution took place—a “proto-music” that would fundamentally implicate music in the development of language and theatre.
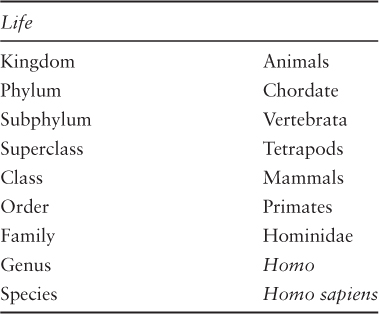
And, oh, by the way, you may not have noticed, but we have slowly been narrowing our scope of inquiry to our human ancestors. Until slugs start writing symphonies or zebras start performing monologues, this seems like a logical choice. Our approach follows what is known in biology circles as taxonomic classification.
We started with the kingdom of Animals, and followed the evolution of our particular phylum, that is, Chordate, and its subphylum, Vertebrata, or vertebrates, animals with a spinal column. We mentioned our superclass, Tetrapods, when they exited the water and started roaming the earth, and out of that lineage, witnessed the evolution of our own class, Mammalia, or mammals. Mammals are animals that have a three-boned middle ear, among other unique characteristics. In this chapter, we will continue to climb the branches of our “Tree of Life,” as Darwin called it. We’ll discover how our order, Primates, further contributed to the evolution of music, language, and the peculiar ability some animals developed to learn the behavior of others called mimesis. We’ll continue to drill down through our family of Hominidae, whose members are called the Great Apes, to our genus, Homo. We’ll explore the precursors to music, language and mimesis, and the earliest evidence of ritual that would ultimately lead to the beginnings of theatre (Darwin 2008).
But before we do, we need to talk for a moment about the many possible ways that evolution may have unfolded. It would be nice if we could simply pronounce one as correct, and follow that one, but at present there simply is not enough evidence to justify that. In fact, there is a strong likelihood that there will never be enough evidence to justify a unified theory of evolution. The fossilized bones and other evidence left behind by our ancestors can only tell us so much. Fortunately, just about every theory leads us to the same conclusion: that music, language and mimesis evolved intermingled. Each phenomenon was intimately involved in the other, and it would have been very hard for one to have evolved without the other. We are, as it turns out, the sum of ourselves, and that remains the whole point of this book: we cannot hope to achieve the best sound designs and compositions for theatre unless we view music as a foundational and holistic part of our theatre experience.
In proceeding in this manner, we will once again run into the common problem: biologists, anthropologists, neuroscientists and others often disagree about the specifics of the evolution of our species. There are those who argue that language either came before music or developed independently of it. I’ll call that the “linguist” approach. It includes Derek Bickerton, who argues that language is based on the ability of Hominins (our human ancestors) to develop symbols, which evolved at a much later date into more complex thoughts using syntax and grammar (Bickerton 1990). This group also includes Steven Pinker, who argues that language is instinctual. Pinker goes on to famously suggest that music is not an evolutionary phenomenon at all, simply “auditory cheesecake,” a bonus that came along with language (Pinker 1997, 524). There are those who argue that language developed from essentially music-like expressions of emotion, especially Darwin (Darwin 1871, 241). There are those who argue that music and language both evolved simultaneously, perhaps out of a common ancestral stage that was neither musical nor linguistic, a “holistic” “musilanguage.” This group includes Steven Brown (Brown 2000), Allison Wray (Wray 1998) and Steven Mithen (Mithen 2006). Finally, neuroanthropologist Merlin Donald proposes that imitation, or its more sophisticated form, mimesis, must necessarily come first in the cognitive evolution of humans (Donald 1993a). And, of course, there are variants on all of these. While acknowledging substantial differences for their purposes, I don’t see these arguments as being contradictory for our purpose: tracing the fundamental role that music plays in the development of what Oscar Brockett calls the autonomous theatre (Brockett 1992, 7).
One would think that there would be more research into the origins of theatre that included its anthropological, biological and neurological origins, but the theories about the origins of theatre tend to focus more on the history of the autonomous theatre, especially Western theatre, the theatre since the Greeks. Since we are interested in how the autonomous theatre developed out of music, how theatre is a type of music, we’ll actually end our journey with the first autonomous theatre in Greece. After all, there hasn’t been much evolution-wise that has changed in the blink of a 2,500-year eye. Fortunately, there are a lot of clues in these evolutionary theories that help us to understand the close and rather intermingled evolution of language, music and mimesis, the primary components of theatre.
Not surprisingly, a lot of the differences of opinion depend quite a bit on how one defines the words “music,” “language” and “mimesis.” We just made such a distinction between music and language in Chapter 4. Hopefully, distinctions such as this will help us not wander too deep into those controversies. There is little controversy about whether language is considered an evolutionary adaptation. David Huron and many others also favor the thinking that music is also an evolutionary adaptation selected to favor survival of the species (Huron 2006, 59). That point of view has perhaps been more definitively supported in the 2015 discovery of specific regions of the brain dedicated to music (Norman-Haignerem, Kanwisher and McDermott 2015). Merlin Donald’s contention that mimesis is an evolutionary adaptation found its neurological evidence in 1992, when a group of Italian researchers discovered the existence of the so-called mirror neuron, perhaps the genetic basis for mimesis (di Pellegrino et al. 1992). We’ll explore all of these discoveries as we move forward in our evolutionary journey.
But rather than spend our precious time debating “who’s on first?,”1 I’ll try to continue on our path of discovering important developments in the evolution of our species. And when we do wander into areas where we are following one researcher’s thesis over another, I’ll always try to make sure that the research in question is cited. I’ll inevitably make no one happy. As Merlin Donald, put it: “Evolution is generally kludge-prone and messy, and deals with terrifying degrees of complexity” (Donald 1993b, 782). But hopefully it will give you a rich set of possibilities to consider. Hopefully it might even influence your aesthetic.
Our journey continues in this chapter as we consider the communicable signs, the music-like expressions, and the emerging imitation in early primates. First, we’ll explore signs, what many suggest are the prototypes of language. Next, we’ll consider the extraordinary evolutionary step our ancestors took as they learned to walk on two legs, an important milestone in our full acquirement of musical rhythm. Finally, we’ll discover a neurological basis for imitation, and consider early manifestations of mimesis, the ability to imitate, rehearse and perform the actions of another. In all three developments, we’ll see the influence of the other two.
Music, Language and Theatre: The Really Early Years
We started this journey 14 billion years ago, and we now find ourselves down to the last 60 million years. And yet it’s hard to imagine a time frame of 60 million years, especially when our recorded history is measured in thousands of years. There is a great tendency to mentally imagine things happening much, much faster than they did. So, when we suggest that about 60 million years ago, our order, primates, first appeared, it’s important to never lose sight that our margin of error can often be much larger than the recorded history of human civilization!
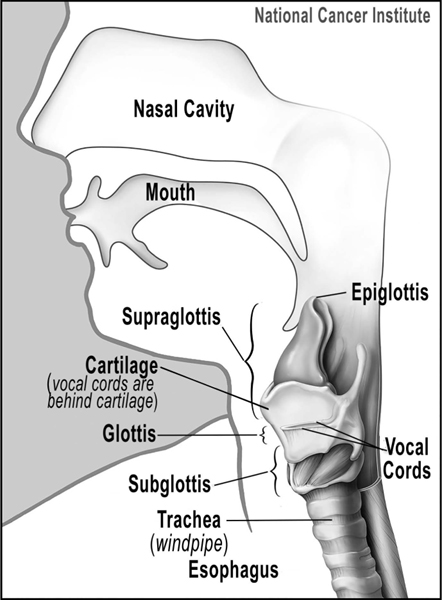
Figure 5.2 The larynx and nearby structures.
Credit: From the National Cancer Institute, Alan Hoofring, Illustrator.
Primates fully developed about 50–55 million years ago, about 10–15 million years after the extinction of the dinosaurs. By this time most members of the mammalian species had developed much more sophisticated vocal tracts that allowed them to produce a wider range of sound colors.
These must have had an evolutionary advantage, as some of these primates grew larger to about the size of a dog, developed larger brains, mainly lived in trees, and tended to be vegetarians (O’Neil 2012). Many mammals that evolved later did not confine their emotional expressions to vocalizations. Deer, rabbits, elephant shrews and skunks, all exhibit a behavior called “foot-drumming,” in which they stomp their feet repeatedly when they encounter a predator or are alarmed (Randall 2001, 1143). The only distant mammal that has showed a capacity for entrainment, the ability to synchronize to a beat, however, is a sea lion named Ronan (Goldman 2013). Hard to say whether there is an entrainment gene lurking somewhere in our distant past, or whether it is an adaptation that has evolved independently from us humans. Part emotion, part sign. At this early proto-stage, it is hard to tell proto-music and proto-language apart.

Figure 5.3 An Old World monkey: mandrill.
Credit: Curioso/Shutterstock.com. Adapted by Richard K. Thomas.
Matt Rossano describes ritual as an attention-getting, sequenced pattern of behavior governed by rules and formality, and meant to convey meaning (2010, 82; 2011, 42). Ritual involves participants performing sequences that do not vary and that the other participants do not create, but instead follow (Smuts and Watanabe 1990, 168). For example, there is the approach ritual of deer, elk and moose, in which the approaching male acts like he’s a calf approaching his prospective mate to nurse, so as not to scare her off (Rossano 2010, 83). This does not surprise me. I have witnessed a fair number of approach rituals by the male species myself in the bars and clubs of college campuses.
Old World monkeys diverged from our line of descent about 30 million years ago2 (Cambridge Encyclopedia of Human Evolution 1992; Nature 2011, 530). They are an important resource about we humans because we both diverged from the same ancestral line. There are two additional traits that we discover in both humans and Old World monkeys that provide clues about music, language and mimesis. The first we’ll talk about is how they express themselves in color and rhythm, both as a primitive form of communication (proto-language), and as a method of expressing emotion (proto-music). The second trait we humans have in common with Old World monkeys is the evolution of imitation, essential to playmaking.
Old World monkeys also exhibit drumming activities in captivity. Of course, even in our more closely related cousins, we need to understand that we are not talking about rock star performances here.
When researchers Remedios and Kayser say that macaque monkeys drum, they mean three things: first, there is a periodic repetition of individual beats; second, these drumming outbursts are short (e.g., they last an average of 1.5 seconds); and third, they are created on “artificial sources,” such as shaken branches or thumped-on logs, so they sound decidedly different than vocalizations. Not exactly Top 40 hit material, but it’s a start when we consider how our inherent sense of rhythm evolved. Researchers often talk about this drumming in terms of expressing emotion (proto-music) or communicating information (a “proto-language,” which is usually accompanied by the “proto-music” of drumming: rhythm, color, etc.). Remedios and Kayser have demonstrated that macaque monkey drumming does activate our old friend, the amygdala, as good an indication there is that drumming is an expression of emotion (2009, 18010–18014).
Research on one species of Old World monkey, the vervet, helps us understand the nature and extent of primitive vocal expression as communication. These monkeys developed distinctly different calls based on the type of predator spotted, so that they could communicate whether the threat was from above (e.g., an eagle), below (e.g., a python), or across (e.g., a leopard), and take appropriate evasive action, such as climbing a tree or hiding in a bush. For example, if the monkeys were on the ground, a leopard call might send them scurrying up trees (Seyfarth, Cheney and Marler 1980). Since the calls themselves indicate specific referents (signs), this type of signaling may represent very early evolutionary evidence of communication using vocalization.
At the same time, it would be hard to argue that the calls themselves did not have a proto-musical component. Imagine the monkey calmly standing by going, “dude, there’s a leopard about to pounce on one of us.” Not likely. Instead, expect a little startle effect, with the monkey barking loudly, perhaps at the top of his or her lungs (Mithen 2006, 108). Such calls would have had distinctive color, rhythm, mass and line characteristics that quickly and effectively changed the ambient emotional environment from calm to mega-tense. Those vocalizations would be accompanied by appropriate physical gestures, perhaps themselves prototypical of music’s visual expression, dance. Darwin famously noted great similarity between monkey facial expressions and humans (Darwin 1871, 133–147). The referential information contained in the vocalization and physicalization certainly would have helped identify the source of the problem and outline a course of action. But the musical aspects would have incited a sense of urgency that would be hard to ignore. The only component it lacked that would qualify it as music would be its artistic function, which, hopefully, was somewhat suppressed at that critical moment. Neurologist Steven Brown describes a signal of this sort as a “referential emotive vocalization” (Brown 2000, 291).


Almost simultaneously in the big picture of evolution, other Old World monkeys including geladas and baboons have been observed in what some anthropologists argue are primitive ritual behaviors.
Geladas developed elaborate social rituals for greeting, departing, grooming and warning each other that involve elaborate singing—modifications of melodic line, rhythm, mass and so forth (Mithen 2006, 109–110).
Baboons greet each other by performing the ritual of scrotum grasping. Yes, it means exactly what you think—male baboons rendering themselves extremely vulnerable while greeting each other.
Living in an environment in which baboons are routinely hostile toward each other, this manner of greeting far surpasses our human ritual of shaking hands, if for no other reason than the obvious potential danger involved (Smuts and Watanabe 1990, 152). Noteworthy is that the scrotum grasping is preceded by lip smacking and other visual gestures to indicate friendly intentions, introducing a proto-musical component into the ritual (Rossano 2013, 96).

Is there a biological basis for such behaviors? It turns out that there may be. In 1992, a group of researchers published startling research that documented the existence of a mirror neuron system in macaque monkeys. While initially controversial, this research seems to be gaining considerable acceptance in the mainstream of science.3 Mirror neurons fire when an action is performed, which is typical for a neuron. But they also fire when an individual observes the same action performed (di Pellegrino et al. 1992). In research experiments, a monkey grasped a cup, and watched a cup being grasped, in both cases firing mirror neurons in their brains. It did not make a difference whether the monkey grasped the cup itself, or watched someone else grasp a cup; whether it was a human or a monkey grasping the cup, whether it was a cup or another object, or how far away the cup was from the monkey. In all cases, the mirror neuron fired. However, if someone just mimed grasping the cup, the neurons would not fire. Mirror neurons appear to be almost exclusively referential, at least in these studies.

The discovery of mirror neurons led to a number of fanciful speculations that really got the subject off to a bad start, leading two of the researchers that discovered mirror neurons, Rizzollatti and Carighero, to remind us that there are two main hypotheses regarding the function of mirror neurons that we must keep in mind. The first is that mirror neurons are an important but not sole part of a primate’s ability to imitate another’s action. There is a lot more going on in the brain when animals, including us, imitate. For example, the mirror neurons have to be programmed or re-programmed in order to function (2004, 172). Catmur, Walsh and Heyes conducted a series of experiments in 2007 that showed that mirror neurons develop through sensorimotor4 learning, sort of a demonstration of a potential underlying mechanism to “monkey see, monkey do” (Catmur, Walsh and Heyes, 2007). The second hypothesis is that mirror neurons serve as the basis for one primate’s understanding of another’s actions (Rizzollati et al. 2004, 172). Mirror neurons do not directly cause imitation, but may provide a key evolutionary adaptation that would allow animals to learn to imitate (Arbib 2013, 118). Of course, it takes a lot more than evolving mirror neurons to make great theatre.
In a 2009 journal article, Marco Iacoboni extended this concept of the relationship between imitation, mirror neurons and understanding to the subject of empathy. Keep in mind that empathy is the ability or tendency to feel what another person is feeling. This differs from sympathy, which is simply the ability to understand what another is feeling, but not to actually feel the same emotion oneself. Iacoboni first observed that authors such as “de Montaigne (1575), Adam Smith (1579), Nietzsche (1881) and Wittgenstein (1980)—have often associated imitation with the ability to empathize and understand other minds” (2009, 654). He describes a “low road” to imitation that “leads to imitation in a direct fashion, such that the perceiver acts the gestures, postures, facial expressions, and speech perceived in other people.” But he goes on to suggest a “high road leads to complex and rather subtle forms of imitation, as shown by a number of experiments that lead to stereotype activation or trait activation” (2009, 657). The “high road” experiments often led to automatic responses of which the participants were not consciously aware. Iacoboni describes experiments specifically tailored to investigating our ability to empathize, that is to feel what someone else is feeling, and imitation. In short, he found that “through imitation and mimicry, we are able to feel what other people feel” (2009, 659). Iacoboni further described experiments that suggested that “the large-scale network composed of mirror neuron areas, insula, and the limbic system (see Figure 5.9) likely provides a simulation-based form of empathy” (665). These of course, are the same areas we have discovered (and will continue to discover!) that music plays an important and often unconscious role in stimulating. Here in a nutshell, we find a neurological and sociological basis for the fundamental processes involved in both music and acting. In later chapters, we will return to this phenomenon, as it has everything to do with the powerful combination of music and mimesis in theatre.
But what about sound and mirror neurons? Can imitating and representing real sounds effect real communication using mirror neurons? In 2002, Evelyne Kholer led a group who investigated whether a monkey’s mirror neurons would fire to the sound of an action all by itself, with no visual reinforcement. In cases where monkeys were familiar with the sound of the action, 13% of the mirror neurons fired even when the visual stimulus was not present. These have been named audiovisual neurons (Kohler et al. 2002). Based on this, the researchers propose that mirror neurons may also be implicated in the development of spoken language as we will investigate in the next chapter.
Credit: Sobotta, Johannes. 1909. Textbook and Atlas of Human Anatomy, edited with additions by McMurrich, J. Volume III. Philadelphia and London, W.B. Saunders. Figure 633, Page 147. Adapted by Richard K. Thomas.
Credit: Blausen.com staff. 2014. “Medical Gallery of Blausen Medical 2014.” WikiJournal of Medicine 1 (2). doi:10.15347/wjm/2014.010. ISSN 2002–4436. CC-BY-3.0. https://creativecommons.org/licenses/by/3.0/deed.en. Adapted by Richard K. Thomas.
Consider the relatively low percentage of audiovisual mirror neurons that fired relative to all of the mirror neurons in the brain.5 We’ve already observed in the last chapter that sound performs rather poorly at “referring” to “real doings.” Perhaps, we may find a neurological basis for this in mirror neurons. But it’s not an entirely visually dominated referential world, either. In the study, there were a few instances where the sound alone stimulated the mirror neurons but the visual stimulus without the sound did not. For example, the cracking of peanuts stimulated the monkey’s audiovisual mirror neurons, but watching someone crack peanuts without hearing the sound did not (Kohler et al. 2002, 847). Some sounds, what I refer to as iconic sounds (sounds that everyone immediately recognizes), carry tremendous referential power. But they tend to be few and far between.
When mirror neurons fire to sounds, meaning becomes attached to elements we describe as proto-music, that is, the color, rhythm, mass, line and so forth of the gesture or sound. These imitative actions, their sounds and finally the names we give them in language seem to be thoroughly intertwined when considered in this manner. It’s hard to even imagine the gestures, music and meaning becoming dissociated from each other, but in modern theatre practice, this is a rather common occurrence.
An old Purdue University production of The Matchmaker required one of my students to create a horse-drawn carriage to pull up offstage. This “effect” was somewhat doomed from the get-go by a number of factors:
- There was no visual gesture (i.e., a real carriage pulling up onstage) to help identify the sound.
- The sound of a carriage moving is not necessarily iconic; it could (and did) sound like a lot of other things.
- The carriage pulls up backstage, and that’s what we hear, a carriage pulling up in the acoustic of the backstage area, not “outdoors” of this “home” as we are led to believe, (the imitation is inherently not very good).
- The sound did not fit into the “music” of the existing scene.
Our poor student sound designer found himself facing what everyone considered to be a simple task (“Just pull the sound out of your sound effects library”) and being the target of reviews like “sounds like a bunch of wood falling.” The solution, of course, would be to attach the sound to a visual gesture, or to referential dialogue (“here comes the carriage!”) that accomplishes the same task. But it is often impossible to do; the carriage is offstage and not visible, the playwright didn’t write the necessary referential dialogue. In these cases, we really, really want the sound to do that referential signaling, just like our monkey friends hearing peanuts being cracked! My friend Joe Pino calls this sort of sound the “offstage one act,” and suggests that he typically recommends truncating such scenes to be as short as possible (Pino 2017). A sound designer caught in the crosshairs of this compromise should first look to more iconic sounds (in this case, for example, lead with a horse whinny). Whether the horse whinnying triggered everyone’s audiovisual mirror neurons as they imagined a carriage arriving offstage is anyone’s guess, but this research certainly helps us to better understand how important it is to approach referential, imitative sound in a play carefully.
Even if we find the perfect iconic horse-drawn carriage sound to use in the scene, our effect may still distract rather than pull us deeper into the scene. The vervet monkey did not stand idly by, nonchalantly signaling the approaching leopard. The vervet monkey call would have been filled with an emotion appropriate to the scene. The fundamental problem related to using sounds from sound effects libraries is that the sound effects were not recorded with the underlying music of the scene in mind. Sticking sound effects on top of the music of a scene is no more useful than putting a party hat on the scrambling vervet monkey. Most of the great sound designers I have ever known disdain using sound effects libraries. They prefer to create each sound through imagining the music of the moment of the scene in which the sound must exist. In other words, they attempt to treat the sound effect as a piece of music, whose instruments are iconic sounds, with each sound orchestrated into the fabric of the scene. In the case of our carriage arriving, a punctuation was required in the rhythmic timing of the actors and the scene, not a long, sustained sound. The music of the carriage needed to not only be matched to the business of the horse, the driver and the carriage itself, but to the music of the scene playing out on the stage in which the audience was hopefully fully engaged. We will talk more about this particular problem in the next chapter when we discuss the concept of entrainment in more detail.
Apes diverged from the same branch of our ancestral tree as monkeys. Gibbons, or “lesser apes” next split off from this branch about 18–20 million years ago6 (Nature 2011).
Gibbons are members of the lesser apes family, the next major branch of our taxonomic classification system. They are physically smaller than the family from which we humans descended, the great apes. Gibbons are particularly known for their beautiful haunting songs in which they produce relatively pure tones in stunning melodies to mate monogamously or to mark their territory. Hiroki Koda and others in Japan discovered that gibbons manipulated their singing voice in the same manner as human opera singers: they “tune” their vocal tract to the fundamental frequency, which amplifies their call considerably, allowing them to be heard from over a mile away7 (2012, 347). This ability of gibbons to manipulate their vocal tracts to produce a wide variety of vocalizations is somewhat unique (musical or linguistic?) in apes. Many apes have obviously not developed vocal abilities to produce such auditory delights. At the same time, gibbons are not known for drumming. But seriously, if you had a voice like that, would you play percussion?
Orangutans, one of the family of great apes that includes us humans, diverged from our human ancestral line about 12–16 million years ago.8
They don’t drum at all, as far as we know, but are said to be great learners, able to learn over 30 unique signs.


Gorillas diverged next, about 6–8 million years ago, although more recent evidence puts the split as early as 8–19 million years ago9 (Nature 2011, 530; Langergraber et al. 2012). They can also learn signs, and like other primates engage in a form of group singing when enjoying certain foods (Shapiro 1982). Notably, they are also avid drummers who like to drum out beats on their chests.
In all of these primates we witness something of a common feature: the animals’ vocalizations indicate and change with the animal’s emotional state. Sometimes the animal is also communicating or attempting to communicate something quite referential, but not necessarily. When they do communicate, they communicate holistically, meaning that they vocalize complete communications that cannot be broken down into syntactical constructions of individual words (Wray 1998, 50–51). Also worth noting and central to our definition of music, expression in apes is multimodal, meaning that it involves gesture and sounds, proto-music and proto-dance. The ape may attach a communication to the qualities we have come to associate with proto-music, but those qualities may also exist simply for their own sake, for example, when gorillas sing while eating certain foods. Lea Leinonen’s research on macaque monkeys showed that even humans can successfully identify their emotional states of fear, aggression, dominance and neutrality, suggesting that we tend to share the same vocalization (i.e., elements of music) patterns (Leinonen et al. 1991). Macaques also tend to lateralize10 musical elements such as color, rhythm and mass in the right brain, and referential qualities in the left brain, as we’ll later discover in humans. So, we are also seeing neurological support that these proto-musical elements are distinct from their proto-language elements, suggesting that, even in the “proto” stage, it is relatively easy for music to exist without language, but rather difficult for any kind of language to exist without music (Hauser 1999, 98). In fact, we won’t see an ability for language to exist independently of music until written language makes its first appearance. We see in all these early primates, then, a rather ubiquitous property of music: it can exist just fine on its own for the purposes of expressing emotion, or it can have referential meanings attached to it that not only communicate ideas, but also incite in the listener emotions similar to the those generated by animal vocalizations and gestures (Hauser 1999, 77–79).
Our closest kin chimpanzees diverged about 4.5–6 million years ago, although, again, new evidences suggest the split may have occurred much earlier, about 7–8 million years ago11 (Nature 2011; Langergraber et al. 2012). Chimpanzees also produce food-associated calls and can learn as many as 250 symbols, taking important evolutionary steps along the way to the emergence of fully developed music, language and imitation in the human species (Luef, Breuer and Pika 2016, 1; Nature 2011; Mithen 2006, 105).


Chimpanzee learning and social behaviors are so divergent and extensive that researchers legitimately refer to them as “cultures.” We have to be careful as we proceed to separate culturally acquired behaviors from evolutionary adaptations, which can be controversial. These behaviors include tool usage, grooming and courtship rituals. The important characteristics of culture are that it is “transmitted repeatedly through social or observational learning,” and that the behaviors exist throughout the community. Separating culture from evolution becomes very important when we think about the communal nature of both music and theatre and our developing sense of rhythm as a social cohesive (Whiten et al. 1999, 682–683; King 2015, 393).
Chimpanzees and their close relatives, bonobos like to drum on resonant objects like tree trunks, and will come back often for a repeat performance when they find one whose sound they like (Center for Academic Research & Training in Anthropogeny 2015). Chimpanzees customarily drum and perform ritual “rain dances” when it starts to rain, further evidence of an intermingling of music, language and mimesis in our distant past.

None of this noise making is particularly synchronized or coordinated in time, however. In Chapter 2, we discovered a number of creatures with an odd ability to synchronize some particular rhythmic activity with each other in a phenomenon called entrainment. But its occurrence in our line of descent, seems somewhat random, and really didn’t appear until much, much later in our evolution. In our own near lineage, we do find what might be called true rhythmic behaviors in bonobos, who exhibit a form of beacon effect called staccato hoot, in which males all bark together in unison to amplify their signal. In both humans and bonobos, the middle range in which human synchrony (entrainment to a beat) is possible is about 2 Hz, or 120 beats per minute. Both humans and bonobos may also have been able to use human movement—akin to bipedalism, but moving in place, as in a dance—to learn to increase the precision of their beat synchronization. Perhaps these commonalities provide an indication that our ability to entrain to beats derived from a common ancestor (Merker, Madison and Eckerdal 2009, 6–7).

Bipedal Primates
After splitting off from chimpanzees, our own ancestors did something quite stunning and remarkable: they stood on two legs and walked upright. Not just some of the time, as some members of our great ape family did, but habitually, a characteristic rare among mammals. Bipedalism evolved somewhere between 6 and 7 million years ago with evidence provided by a closely related genus, Sahelanthropus.
Genus is the next major narrowing of our taxonomic classifications after family. Sahelanthropus had a chimpanzee size brain, but the foramen magnum, the hole in its skull where the spinal cord enters the cranium, was at the bottom of the skull, as in humans, rather than in the rear, as in other apes (Brunet 2002, 2005).
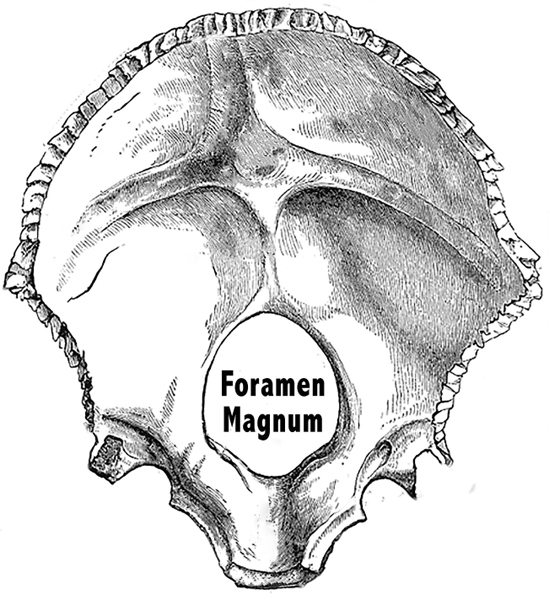
Figure 5.16 The foramen magnum, at the base of the human skull, allows humans to walk upright.
Credit: Gray, Henry. 1918. Anatomy of the Human Body. Lea and Febiger. Plate 130. Adapted by Richard K. Thomas.
This led anthropologists to conclude that Sahelanthropus was the first member of the Hominini tribe to walk upright. Such a restructuring of the skull would also have effected changes to the vocal tract, beginning a long pattern of evolution that would eventually see the larynx descend further down the throat (Mithen 2006, 129).
Cognitive scientist Philip Lieberman proposed that the evolution of human walking provides evidence of an evolving basal ganglia. The basal ganglia consist of groups of neurons that sit at the base of the cerebral cortex (the outer layers of the brain) indicating that the basal ganglia are a very primitive and old part of our brain. They are associated with controlling voluntary motor functions, learning and emotion.
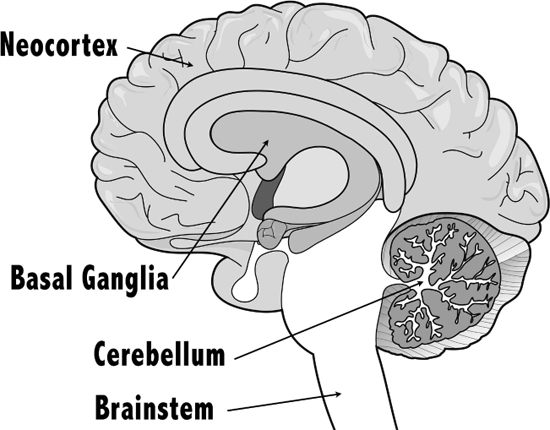
Figure 5.17 The basal ganglia and related structures.
Credit: Blamb/Shutterstock.com. Adapted by Richard K. Thomas.
The basal ganglia work in conjunction with the cerebellum, the oldest part of our brain, and our new brain, the neocortex, to “play a critical role in motor control, motor learning and cognition.” Both walking and speech require the basal ganglia to work as a “‘sequencing engine’ that can reiterate motor commands stored as motor pattern generators in other parts of the brain,” putting together sequences of instructions into meaningful actions. Lieberman suggests that walking was perhaps the “starting point” for the evolution of human speech, language and cognition (Lieberman 2007, 47–52).
Steven Mithen argues that the basal ganglia are also the starting point for our superb sense of rhythm, as rhythm is essential to walking on two legs (Mithen 2006, 150). There is a direct relationship between music and walking in the basal ganglia: rhythm. The basal ganglia must jump into action anytime an internal beat must be generated, and this includes both walking and playing music with a beat. Chimpanzees and apes are bipedal for short amounts of time, but their ability to generate and maintain a beat is not ubiquitous or customary in nature, and very difficult for them to acquire in captivity. In Sahelanthropus and their descendants over the next few million years, we see a possible origin that connects the act of walking and our ability to generate and maintain a beat in musical rhythm.
About 4 million years ago, another genus, Australopithecus, emerged, and it has a very popular representative in Lucy, a 3.2-million-year-old skeleton, who captured the imagination of the world when she was first discovered in 1974 (Institute of Human Origins 2014; Martin 2015, 38; Falk 1999, 211).

Figure 5.18 Australopithecus reconstruction.
Credit: Nicolas Primola/Shutterstock.com. Adapted by Richard K. Thomas.
And while Lucy may have walked, Steven Mithen points out that she was not the first to run and jump, as it would take another million and a half years before bipedalism would develop to the full capabilities humans enjoy today (Mithen 2006, 139–144).
Bipedalism did allow Australopithecines to wander out of the forests and onto the savannahs. Mithen argues that this new habitat pressured Australopithecines to develop a more sophisticated and wider range of calls than apes or monkeys. They developed an ability to alert the presence of a group of predators or found food that would best be shared in an expanding social environment. Musically, Mithen suggests that Australopithecines developed the ability to modulate the mass (volume) of their calls to signal without alerting predators (“Shhh … there’s a lion over there!”), and much more musically expressive rhythms and lines (melodies) than their gelada ancestors. But Mithen also tells of one archaeological find, a child swooped away from its mother by an eagle, that perhaps elicited the most musical vocal expression of all: the lament of a mother over a lost child (2006, 132). Neuroanthropologist Dean Falk concludes that these early ancestors used vocalization especially to express emotions. Falk based his analysis on chimpanzee social and vocalization habits, as well as their vocal tracts and cerebral cortices, emphasizing the more music-like purpose of their vocalizations over the referential (Falk 1999, 211). Mithen summarizes the communications of these early Hominins12 as “Hmmmm:” Holistic (meaning that each vocalization comprised a complete thought rather than a part of a thought such as a word), multimodal (typically involving vocalization and gesturing), manipulative (a command or a request rather than an abstract thought) and musical (involving manipulations of melodic line, mass, rhythm, etc.) (2006, 138).
About three and a half million years ago, early Hominins13 roamed eastern and southern Africa, and developed a very primitive ability to make rudimentary stone tools.
These tools are called Oldowan, after the lake in Tanzania where they were first discovered. They were basically chopping tools used for, among other things, cutting the flesh off of slaughtered animals (Mithen 2006, 125). What makes these tools so special is that it took a certain amount of sophistication to make them; in order to get their sharp edges, they had to be fashioned by striking one stone against another in a certain way to get a sharp edge, and to flake off other bits of stone that could be used for other “tool” purposes.
Merlin Donald sees in this tool-making process one of the first clues to developing human cognition. Donald thinks that Australopithecines might have been the first to break two mental barriers: first, the ability to voluntarily recall memories, and second, the ability to express knowledge in some sort of action, even, as he says “in simple pantomime”14 (Donald 1993b, 35). The ability to voluntarily recall memories would allow the Australopithecine to voluntarily recall a technique that worked in creating a tool, to demonstrate that technique to another, for the other Australopithecine to imitate it, and then to rehearse the learned technique until perfected (Donald 2012, 740–741). What a significant advantage this would have provided Australopithecines over other animals! Donald calls this ability mimesis, and expressly understands it to be the same basis for our much later cultural forms that include theatre and dance (Donald 1993a, 153).

Credit: Reprinted by permission from Macmillan Publishers Ltd: Nature. Hovers, Erella. “Archaeology: Tools Go Back in Time,” Nature 521, 294–295. doi:10.1038/521294a, copyright 2015.

Nevertheless, despite having improved on earlier hominid ability to develop and use tools, Australopithecines were not considered to be all that smart. Their brain size was only slightly larger than chimpanzees, weighing in at around 400–500 cubic centimeters (Mithen 2006, 123). While Donald suggested that the roots of mimesis lay in Australopithecines, he nevertheless concluded that “they did not leave any evidence suggesting major cognitive evolution” (Donald 1993b, 738). The next major chapter of cognitive development would be evidenced in a genus much closer to us anatomically and functionally, Homo. We’ll explore them, and their contributions to music, language, and theatre in the next chapter.
Ten Questions
- Trace the taxonomic classification of our human ancestors from kingdom through genus.
- What two types of rhythmic behaviors do we witness in early mammals, and which animals exhibit them?
- Name two significant traits we find in both humans and Old World monkeys.
- Define ritual and give a mammalian example.
- What is uniquely different about mirror neurons from other neurons?
- What do the basal ganglia and the foramen magnum have to do with Sahelanthropus?
- What is the biggest difference between chimpanzee and human drumming, and to what evolutionary development do some researchers ascribe that difference?
- What in the world do mirror neurons have to do with choosing iconic sounds in theatre sound design?
- The elements of color, mass, rhythm and line have been observed to be used in two ways by apes. What are they?
- Why does Merlin Donald think that the ability of Australopithecines to make tools is so important?
Things to Share
- In the last chapter, you explored manipulating color to incite various emotions in your audience. Since Australopithecines became more proficient in manipulating the mass (volume, shape, dynamics) of their vocalizations, we’ll also explore manipulating intensity to stir emotions in your audience. To do this, we’ll want to hold color constant, and let it have as little influence as possible on our audience. In this game, you get to use one breath, in which you may vocalize the “Shhhhhhhhh” sound for as long as you care in order to elicit each of the following emotions in your audience: love, anger, fear, joy, sadness (i.e., one breath per emotion!). Be careful not to manipulate the color of the sound by changing the formants in your throat, etc. Instead just manipulate the loudness of the sound to create five samples, one for each emotion. Record these five sounds as five different audio files, and label them “Take 1, Take 2,” etc.
We will play your five sounds back for the class, and the class will have to suggest which examples most incited which emotions in them. We’ll poll the class to discover which sounds they associated with each emotion, and then we’ll ask you for the order you intended. We’ll then tabulate the percentage of emotions you were able to correctly incite in each audience member, and discuss the results. - Having explored the wonderful world of color, now it’s time for you to explore color in composition. Compose or design a 1:00–1:30 piece that does not use a well-defined tempo, but instead, focuses on the interplay of a plethora of sonic colors. Work to stimulate a strong emotional response in your audience without using referential, iconic sounds—or at least without using them to reference their original source! (Feel free to manipulate and mangle well known sounds to create emotional responses, however.) Use original sounds that you have gone out and recorded yourself. Shun the sound effects libraries! Avoid traditional musical instruments such as guitar or piano! Find new sounds in the real world; long, sustained sounds, quick transient sounds, and everything in between. Sample those sounds and then manipulate and twist them into something entirely new! Orchestrate many sounds into your piece rather than just a few. Play and experiment!
Notes
1For our younger readers, “Who’s on First?” is a famous vaudeville routine performed by the comedy duo of Abbott and Costello in the 1940s. It’s readily available for viewing on the internet.
2The branch that includes both monkeys and apes is called a Parvorder, and the classification is Catarrhini. You will notice that I’m putting a lot of these more minor taxonomic classifications in footnotes, as they can get quite confusing, and I’m absolutely certain that anyone can become a great composer and sound designer without having mastered them. Still they are informative and interesting, so I don’t want to leave them out.
3For example, see the feature on the popular PBS series, NOVA (2005).
4Quite literally the combination of sensory systems and motor systems in the brain.
5In the particular area of the brain observed, F5 of the premotor cortex. See Figure 6.9.
6In our taxonomy, apes belong to the superfamily Hominoidae, that also includes humans.
7There are many videos on YouTube of gibbons singing. Look for ones in their native lands of Laos or Cambodia, where the natural acoustics of the forests add immensely to the beauty of their songs.
8The technical name for this family is Hominidae—not to be confused with our superfamily Hominoidae!
9In the subfamily Homininae—is this getting complicated or what? Fortunately, you don’t really need to know all this in order to be a great composer for theatre! But it does help to follow the evolutionary chain that helps us understand how we came to be so musical.
10Just a fancy way of saying “left brain” or “right brain.”
11In the tribe Hominini.
12Hominins are a subtribe of Hominidae, that only include our human ancestors, not the other great apes.
13We say Hominin here, because there isn’t general agreement on whether Australopithecines or early Homo created these tools, and they are both Hominins.
14Note that Donald first proposed these characteristics in Homo erectus in 1993, but updated to include some rudimentary form of it in the earlier Australopithecines due to their ability to make tools. See Donald (2012, p. 30).
Bibliography
Arbib, Michael, A. 2013. “Précis of How the Brain Got Language: The Mirror System Hypothesis.” Language and Cognition 5 (2): 107–131.
Bickerton, Derek. 1990. Language and Species. Chicago: Chicago University Press.
Brockett, Oscar G., and Franklin J. Hildy. 1992. History of Theatre. Boston: Allyn and Bacon.
Brown, Steven. 2000. “The ‘Musilanguage’ model of music evolution.” In The Origins of Music, edited by Nils L. Wallin, Björn Merker, and Steven Brown, 271–300. Cambridge, MA: MIT Press.
Brunet, Michel, and others. 1992. “Cambridge Encyclopedia of Human Evolution.” In Evolution of Old World Monkeys, edited by J.S. Jones, R.D. Martin, D. Pilbeam and Sarah Bunney, 217–222. Cambridge: Cambridge University Press.
———. 2002. “A New Hominid From the Upper Miocene of Chad, Central Africa.” Nature 418 (6894): 145–151.
———. 2005. “New Material of the Earliest Hominid From the Upper Miocene of Chad.” Nature 434: 752–755.
Catmur, Caroline, Vincent Walsh, and Cecilia Heyes. 2007. “Sensorimotor Learning Configures the Human Mirror System.” Current Biology 17: 1527-1531.
Center for Academic Research & Training in Anthropogeny. 2015. “Drumming.” Accessed March 9, 2016. https://carta.anthropogeny.org/moca/topics/drumming.
Darwin, Charles. 1871. The Descent of Man. London: John Murray.
———. 2008. “The Origin of the Species.” August. Accessed March 7, 2016. http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=text&pageseq=133.
di Pellegrino, G., L. Fadiga, L. Fogassi, V. Gallese, and G. Rizzolatti. 1992. “Understanding Motor Events: A Neurophysiological Study.” Experimental Brain Research 91 (1): 176–180.
Donald, Merlin. 1993a. “Human Cognitive Evolution.” Social Research 60 (1): 143–170.
———. 1993b. “Précis of Origins of the Modern Mind.” Behavioral and Brain Sciences 16 (4): 737–791.
———. 2012. “Northrop Frye and Theories of Human Nature.” University of Toronto Quarterly 81 (1): 29–36.
Falk, Dean. 1999. “Hominid Brain Evolution and the Origins of Music.” In The Origins of Music, edited by Nils L. Wallin, Bjorn Merker, and Steven Brown. Cambridge, MA: MIT Press.
Goldman, Jason G. 2013. “Ronan the Sea Lion Dances to the Backstreet Boys. So What?” April 4. Accessed March 14, 2016. http://blogs.scientificamerican.com/thoughtful-animal/ronan-the-sea-lion-dances-to-the-backstreet-boys-so-what/.
Hauser, Marc D. 1999. “The Sound and the Fury: Primate Vocalizations as Reflections of Emotion and Thought.” In The Origins of Music, edited by Nils L. Wallin, Björn Merker, and Steven Brown, 77–102. Cambridge, MA: MIT Press.
Huron, David. 2006. “Is Music an Evolutionary Adaptation?” Annals of the New York Academy of Sciences 930: 43–61. doi:10.1111/j.1749-6632.2001.tb05724.x
Iacoboni, Marco. 2009. “Imitation, Empathy, and Mirror Neurons.” Annual Review of Psychology 60: 653–670.
Institute of Human Origins. 2014. “Lucy’s Story.” Accessed March 8, 2016. https://iho.asu.edu/about/lucys-story.
King, Glenn E. 2015. Primate Behavior and Human Origins. London: Routledge.
Koda, Hiroki, Takesha Nishimura, Isao T. Tokuda, Chisako Oyakawa, Toshikuni Nihonmatsu, and Nobuo Masataka. 2012. “Soprano Singing in Gibbons.” American Journal of Physical Anthropology 149 (3): 347–355.
Koelsch, Stefan. 2014. “Brain Correlates of Music-Evoked Emotions.” Nature Reviews Neuroscience 15 (3): 170–180.
Kohler, Evelyne, Christian Keysers, M. Allessandra Umiltà, Leonardo Fogassi, Vittorio Gallese, and Giacomo Rizzolatti. 2002. “Hearing Sounds, Understanding Actions: Action Representation in Mirror Neurons.” Science 297 (5582): 846–848.
Langergraber, Kevin E., Kay Prufer, Carolyn Rowney, Christophe Boesch, Catherine Crockford, Katie Fawcett, Eiji Inoue, and others. 2012. “Generation Times in Wild Chimpanzees and Gorillas Suggest Earlier Divergence Times in Great Ape and Human Evolution.” Proceedings of the National Academy of Sciences 109 (39): 15716–15721.
Leinonen, Lea, Ilkka Linnankoski, Maija-Liisa Laakso, and Reijo Aulanko. 1991. “Vocal Communication Between Species: Man and Macaque.” Language & Communication 11 (4): 241–262.
Lieberman, Philip. 2007. “The Evolution of Human Speech, Its Anatomical and Neural Bases.” Current Anthropology 48 (1): 39–66.
Locke, Devin P., et al. 2011. “Comparative and Demographic Analysis of Orangutan Genomes.” Nature. January 26. Accessed March 8, 2016. www.nature.com/nature/journal/v469/n7331/full/nature09687.html.
Luef, Eva Maria, Thomas Breuer, and Simone Pika. 2016. “Food-Associated Calling in Gorillas (Gorilla g. gorilla) in the Wild.” PLOS ONE 11 (2): 16.
Martin, Robert D. 2015. “Primate Evolution.” In Basics in Human Evolution, edited by Michael P. Muehlenbein, 31–41. San Diego: Elsevier.
Merker, Bjorn H., Guy S. Madison, and Patricia Eckerdal. 2009. “On the Role and Origin of Isochrony in Human Rhythmic Entrainment.” Cortex 45 (1): 4–17.
Mithen, Steven. 2006. The Singing Neanderthals. Cambridge, MA: Harvard University Press.
Norman-Haignerem, Sam, Nancy G. Kanwisher, and Josh H. McDermott. 2015. “Distinct Cortical Pathways for Music and Speech Revealed by Hypothesis-Free Voxel Decomposition.” Neuron 88 (6): 1281–1296.
O’Neil, Dennis. 2012. “The First Primates.” Accessed March 7, 2016. http://anthro.palomar.edu/earlyprimates/early_2.htm.
Pinker, Steven. 1997. How the Mind Works. New York: Norton.
Pino, Joe, interview by Rick Thomas. 2017. Personal Interview, April 21.
Randall, Jan A. 2001. “Evolution and Function of Drumming as Communication in Mammals.” American Zoologist 41 (5): 1143–1156.
Remedios, Ryan, Nikos K. Logothetis, and Christoph Kayser. 2009. “Monkey Drumming Reveals Common Networks for Perceiving Vocal and Nonvocal Communication Sounds.” Proceedings of the National Academy of Sciences of the United States of America 106 (42): 18010–18015.
Rizzolatti, Giacomo, and Laila Craighero. 2004. “The Mirror-Neuron System.” Annual Review of Neuroscience 27: 169–192.
Rossano, Matthew J. 2010. Supernatural Selection: How Religion Evolved. New York: Oxford University Press.
———. 2011. “Setting Our Own Terms: How We Used Ritual to Become Human.” In Neuroscience, Consciousness and Spirituality, edited by Harald Walach, Stefan Schmidt, and Wayne B. Jonas. New York: Springer.
———. 2013. Mortal Rituals. New York: Columbia University Press.
Seyfarth, Robert M., Dorothy L. Cheney, and Peter Marler. 1980. “Monkey Responses to Three Different Alarm Calls: Evidence of Predator Classification and Semantic Communication.” Science 210 (4471): 801–803.
Shapiro, Gary L. 1982. “Sign Acquisition in a Home-Reared/Free-Ranging Orangutan: Comparisons With Other Singing Apes.” American Journal of Primatology 3: 121–129.
Smuts, Barbara B., and John M. Watanabe. 1990. “Social Relationships and Ritualized Greetings in Adult Male Baboons (Papio Cynocephalus Anubis).” International Journal of Primatology 11 (2): 147–172.
Whiten, A., J. Goodall, W.C. McGrew, T. Nishida, V. Reynolds, Y. Sugiyama, C.E.G. Tutin, R.W. Wrangham, and C. Boesch. 1999. “Cultures in Chimpanzees.” Nature 399: 682–685.
Wray, Allison. 1998. “Protolanguage as a Holistic System for Social Interaction.” Language and Communication 18 (1): 47-67.