
Genetic Algorithm Optimisation of Fish Shape and Swim Mode in Fully-Resolved Flow Field
Sho Kusudaa; Shintaro Takeuchia; Takeo Kajishimaa a Dept. of Mechanical Engineering, Osaka University. 2-1 Yamada-oka. Suita-city. Osaka 565-0871. Japan
Publisher Summary
Finned aquatic animals, such as fish and dolphin, often exhibit extraordinary physical capabilities for swimming speed. However, it is impossible to manufacture and test all possible types of robot to search good performance because one robot fish has many adjustable parameters for identifying shape and swim mode. The shape and swim mode optimized with energy show qualitative agreement with the theoretical and experimental predictions reported in the literatures. Also, parallel processing is attempted to the GA optimization, and the approach is shown to exhibit a remarkable applicability for large-scale parallel computation. The observation suggests that the fish with non-zero wave number is more efficient in converting the hydrodynamic force into propulsion, while the fish with little phase-lag effectively transfers kick power to fluid to get strong thrust. The flow field was fully resolved from the boundary layer to the far field of the object by time-dependent, body-fitted coordinate system. The authors are currently testing more realistic fish models with increased number of characteristic parameters. A series of new attempts and results will be presented as a separate paper in a relevant journal.
Flow field around a single 2-D fish is analysed by solving the full Navier-Stokes equations. and optimisation of fish shape and swim mode with genetic algorithm (GA) is attempted. To solve the flow field around the moving fish meshed with time-dependent body-fitted coordinate system, arbitrary Lagrangian Eulerian (ALE) method is employed. A study with specific parameters shows that swim mode with/without phase-lag causes distinct differences in fluid’s velocity and pressure fields past a fish, suggesting that the phase-lag parameter plays an important role in the interaction between the fluid and fish’s global movement. For pursuing good performances of fish, five characteristic parameters are optimised in conjunction with the surrounding fully resolved flow field. The GA results show that fish exhibits different ideal shapes and manoeuvres for different desired fitnesses. Fish of efficient energy consumption qualitatively agrees with the prediction by Lighthill[l]. On the other hand the best-performing fish attaining the furthest distance in a given time exhibits totally opposite trends for shape and swim mode with little consideration on efficiency. The study shows that difference in energy consumption for those two extreme cases is about 15 times.
1 INTRODUCTION
Finned aquatic animals, like fish and dolphin, often exhibit extraordinary physical capabilities for swimming speed, efficiency or steep turn, and the performances have encouraged a number of scientists to study the potential capacities of them from many aspects. Meanwhile, ship and undersea vessel have been using rotary propeller to get thrust. If the above fish-like propulsion system is applied to man-made vessels, it may drastically improve efficiency or noise level. In reality, several prototypes of robot fish have been developed already, and in the future the competition in development and improvement of robot fish will be further accelerated. However, it is impossible to manufacture and test all possible types of robot to search good performance, because one robot fish has many adjustable parameters for identifying shape and swim mode.
Therefore, a sophisticated optimisation method is needed to seek superior samples efficiently. There is one particular technique for optimisation, genetic algorithm (GA). suitable for problems of multi-dimensional multi-peaked solutions. GA has learnt its basic idea. i.e. reproduction, crossover and mutation, from the natural evolution system of individuals in a group of families, and it. can be easily modified to search optimal solution through a various situations. Due to its controlled non-linear stochastic process, GA solves the optimisation problem efficiently, and it has gained popularity for predicting optimal solution and efficient machine learning process. For robot fish, Barrett[2] conducted some experiments to optimise fish locomotion by directly incorporating the experimental results into GA, and reported that GA works effectively to achieve desired robot-fish functions. Kuo and Grierson[3] employed GA to search efficient prospective gait based on computational results. Five parameters characterising the carangiform locomotion were optimised in their work. However, their model for fluid-object interaction was rather simplified and restricted by employing an empirical fluid drag model. As object’s global motion is affected by fluid motion passing by it and vortical structures in the wake[4], fully-resolved flow field is of importance in predicting good performance of fish by numerical optimisation.
Due to the recent increase in computational power, large scale numerical simulations of fluid dynamics are carried out. Generally, numerical simulation of fluid-solid interaction needs treatments of both fluid and solid phases in a coupled fashion. Arbitrary Lagrangian-Eulerian (ALE) method is one of the approaches to solve the interaction problems of fluid and deformable object [5]. The method resolves a flow field at a desired scale along object surface, and has found applications in a number of flow fields accommodating moving object. For example, Liu et al[6] applied their finite volume ALE approach to a problem of tadpole swimming, and studied the effect of morphological development of the tadpole on the hydrodynamic forces and efficiency.
In the present work, interaction between a simplified model fish and fluid flow is studied. The fish locomotion in a fully-resolved flow field is facilitated by our ALE method efficiently combined with a mesh generation procedure. GA optimisation is applied for five parameters characterising the fish’s shape and swim mode, and the effect of the parameters on the fish movement and the flow field are discussed. Also, parallel processing is attempted to the GA optimisation, and our approach is shown to exhibit a remarkable applicability for the large scale parallel computation.
2 GOVERNING EQUATIONS AND NUMERICAL METHODS
2.1 Fluid phase
Body-fitted curvilinear coordinate system is employed for fluid phase. In the present study, the coordinates alter to fit the object surface which changes the shape with time in a prescribed manner.
Governing equation of fluid motion is Navier-Stokes (N-S) equation of ALE form[5] and the equation of continuity for incompressible fluid, which are described as follows:
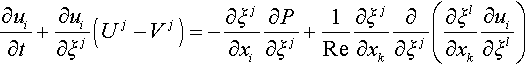

where ui is velocity, p pressure. Re Reynolds number. xj Cartesian coordinates, ξj curvilinear coordinates, Uj and Vj contravariant velocities of fluid and grid, respectively, and J is Jacobian of coordinate transformation between xj and ξJ.
2.2 Solid phase
In the present study, the authors employ a simplified 2-D fish model with a limited number of parameters to easily isolate the effect of each parameter (from the others) on fish performance and flow structure. The fish is allowed three fundamental degrees of freedom (two for translating motion and one for rotating motion) and some additional degrees for the joints on the vertebra. The three fundamental degrees of freedom represents the global behaviour of the fish movement. and they are governed by the following equations:

where m is mass. ![]() velocity of gravity centre. Fi fluid force, dΩ surface element of the object. Li angular momentum. Mi first order moment of the fluid force.
velocity of gravity centre. Fi fluid force, dΩ surface element of the object. Li angular momentum. Mi first order moment of the fluid force.
A NACA0012 hydrofoil is used here as a fish model at the neutral position. Five characteristic parameters are considered to identify individuals. Figure 1(a) shows a schematic of the neutral and deformed positions of the fish and the symbols for the parameters. Within the entire body length L, the fish is assumed to have a deformable rear part (length lp) and a rigid front part (L−lp). The thickness h at the neutral position is 0.12L. In the present study, amplitude a is selected in a range including 0.2L, which is reported as a typical amplitude of a real fish’s movement by Bainbridge[7]. Wavenumber k of undulatory motion (Fig. 1(a). right) is tested in the range between 0 and 0.4. Here. zero wavenumber means that the everv point of the backbone is set in phase (in-phase motion), and non-zero wavenumber causes some phase-lag between arbitrary two points on the deformable: part (phase-lag motion). For both extreme vales of k, volume deviations during the swim are found to be less than 10−5 with respect to the initial value. Frequeney f determines the speed of the undulatory motion propagating down towards the tail tip as 2π f/k. The all five parameters are kept constant for each fish, except that amplitude a is gradually increased in the first one unit time to avoid unnecessary numerical instability. Density ratio of fish to fluid is set to 1.0.

2.3 Numerical and boundary conditions
In the present study, fish is set in an open space. C-type mesh system is used for the simulation of the flow field. Figure 1(b) shows a mesh near the object, and Fig. 1(c) outlines the computational domain and boundary conditions. The computational domain is a semi-circle of diameter 8L in the front of the hydrofoil and 7L in length in the behind. Boundary condition of fluid velocity is the non-slip condition at the surface of the fish (N). At the inlet (S) and out-flowing boundaries (C), stationary fluid and convective boundary condition are assumed, respectively. Neumann’s condition is applied for pressure at the inlet boundary. A traction-free boundary condition is employed at the upper and lower straight sections of the domain (T).
The number of grid points is 240×40. There are 120 grid points on the fish surface. A computational mesh fitted to a deformed fish is given every time step by the elliptic mesh generation algorithm.
3 SPECIFIC STUDY OF PROPULSIVE STAGE
Some specific cases of typical fish characteristics are studied to investigate the fundamental dynamics of fluid and the fish-like deformable hydrofoil. In this section, a, lp and f are set to 0.2L, 0.8L and 1, respectively. Two wavenumbers k=0 and 0.4 are tested to compare the fluid behaviours past the object.
Figures 2(a) and 2(b) compare diagrams of fish velocities and instantaneous flow fields in the initial propulsive stage for the different k values. From the velocity diagrams, the fishes experience large accelerations in both x and y directions after t=0.5. Then, for both k values, x components of velocity develop exponentially with small fluctuations, which eventually attain stable oscillations around the constant cruising velocities at about t=20. On the other hand, rather stable oscillations are observed for the y components around small non-zero constant y velocities, suggesting that the fishes travel in specific directions.

The above fish motions may be related to the fluid motions. Figures 2(d) and 2(e) compare instantaneous pressure and velocity fields in the slip streams for k=0 and 0.4, respectively. The corresponding moments are depicted in Fig. 2(a) as points A and B. In both snapshots, fish tail is in the movement towards the largest displacement in −y direction, generating a vortex of positive circulation in the upper side and a high pressure region in the lower side of the tail.
The difference in wavenumbor has caused a formation of stronger vortex in the slip stream of k=0 case (Fig. 2(e)) than k:=0.4 (Fig. 2(d)). Generally, fish’s total gain in momentum is dependent on the momentums held by the discharged vortices. Therefore. terminal travelling velocity of fish is dominated by the strength of the series of vortices. This may explain the difference in terminal velocities observed in Fig. 2(a).
However, the fish of phase-lag motion overcomes this weakness of generated vortices by its efficient, swim mode. The phase-lag fish develops a positive pressure gradient across the convex and concave sides of the trailing edge, as compared in Fig. 3(a)(b) for the respective instances of Fig. 2(c)(d). This adverse pressure gradient helps the phase-lag fish to be less decelerated in each fluctuating cycle and over the initial propulsive stage (Fig. 2(a)). and consequently, allows the fish to swim efficiently at large. The vortex dynamics induced by the efficient swim mode is the subject of ongoing research by the present authors.

The above observation suggests that the fish with non-zero wavenumbor is more efficient in converting the hydrodynamic force into propulsion, while the fish with little phase-lag effectively transfer kick power to fluid to get strong thrust.
Also the results demonstrates that only fully-resolved flow field enables analysis and prediction of fish-fluid dynamics at this detailed level.
4 OPTIMISATION WITH GENETIC ALGORITHM
It would be interesting to ask what fish shape and swim mode give optimal solution for a given objective. For this question, a genetic algorithm (GA) is employed in the present study, and fish’s characteristics and performances optimised for travelling distance and efficiency are compared.
4.1 Fitness functions
The following five parameters of fish is varied in search for optimised performance of fish: a, lp, h, f and k. Each parameter has 16 equally-divided quanta in the respective range, shown in Table 1. including the values studied in the previous section.
Table 1
Ranges of the GA parameters
| Parameter | h | a | f | lp | k |
| Minimum | 0.08L | 0.08L | 0.60 | 0.5L | 0 |
| Maximum | 0.23L | 0.23L | 1.35 | 1.0L | 0.4 |

To qualify the adaptivity of the individual fish to the environment, the present study employs the following two fitness functions: distance travelled in the first 10 unit time and efficiency (travel distance per unit energy input) in the same duration of time. The fitnesses functions, fd and fe, arc defined as follows:

where GAd and GAe are GA session names targeting optimisation of fd and fe, respectively. For both fitnesses, higher value means better performance. The conditions used for both sessions are summarised in Table 2.
4.2 Results and discussion
Figure 4 compares the progresses in distance and efficiency of GAd and GAe. Figure 4(a) shows that GAd acquires a remarkable capability of long range travel as generation goes, paying little attention to the optimisation of efficiency. Similarly, GAe improves the efficiency progressively despite the poor travel distances. The ratio of energy consumptions of GAd and GAe at 30th generation is about 15 times. This result suggests that an individual optimised in one environment is not always suited for another environment.

Trends of development of each parameter are shown in Figure 5. Experimental results of a real mackerel by Bainbridge[7] and predictions by Lighthill[1] are also included for some parameters. Significant differences are observed between the trends of the solutions of GAd and GAe.

The overall trend for GAd session is summarised as follows. Amplitude, deformable body length and frequency are optimised to near-maximum values of the respective parameter ranges as shown in Figs. 5(a), 5(b) and 5(d). Also, from Fig. 5(e), smaller wavenumber is suitable for the purpose, which is in agreement with the result of the in-phase motion presented in the previous section. The result suggests that, to attain a longer distance in a certain duration of time, the fish twists its entire body as quickly as possible with a large amplitude and little phase-lag within the body.
On the other hand, the optimised fish in GAe has totally different traits; a small amplitude, a shortest possible deformable section, a very thin body, and a largest possible phase-lag but a slow wave propagation. The fish characteristics predicted in GAe agree with the prediction by Lighthill[1] for real fish, and this result may point to the fact that real fish give first priority to efficiency, not to travel distance or speed, in a usual situation.
4.3 Optimisation with multiple processors
Genetic algorithm requires a long run over generations. Particularly, the present case involves fully-resolved numerical simulation of fluid phase, and computational time for the whole process becomes a serious issue. For this problem, we attempted a parallelised processing within each generation by the message passing interface (MPI) library employing 16 processors at. the maximum. The computer used is Dell Precision 350 with processor of 2.66 GHz Pentium4 inter-connected by 5224 Gigabit switches. The trend of computational time for one generation is plotted in Fig. 6. The elapsed time is inversely proportional to the number of processors, suggesting that the MPI parallel computation is suited for this type of problem.

5 CONCLUDING REMARKS
Two dimensional simulation of flow field past a fish-like deformablc hydrofoil was carried out with arbitrary Lagrangian-Eulerian method. The flow field was fully resolved from the boundary layer to the far-field of the object by time-dependent body-fitted coordinate system. Also fish shape and swim mode are optimised by genetic algorithm (GA).
Our preliminary study on fluid-fish interaction showed that the flow structures in the fish’s near and far fields are dependent on wavenumber of the undulatory motion of the fish and that the swim modes affect the hydrodynamic forces acting on the fish surface and terminal velocity of the fish.
It was found that GA solution for one objective is not always suited for another objective. The GA result showed that the fish attaining the furthest range in a given time exhibits totally opposite trends for shape and swim mode to the least energy-consuming fish in unit travel distance. The shape and swim mode optimised with energy show qualitative agreement with the theoretical and experimental predictions reported in the literatures.
And in the present study, our method demonstrated the capability of solving this type of multi-objcctive-multi-solution problem by including the fully-resolved flow field surrounding the object. And the computation time for the large scale GA was found to be effectively reduced by parallel computation.
The authors are currently testing more realistic fish models with an increased number of characteristic parameters (e.g. adaptive undulatory motion) and aiming for further improvement of fish performance with an original acceleration technique of GA. A series of new attempts and results will be presented as a separate paper in a relevant journal.
ACKNOWLEDGEMENT
The authors gratefully acknowledge the Merit Allocation Scheme Grant of the Australian Partnership for Advanced Computing (APAC).