
Chapter 6
Campfire Songs
Rhythm and Entrainment
Introduction: Welcome Homo
In the spring of 1990, the chair of our department, Jim O’Connor, gradually became more aware that my interest in theatre was not simply in the mechanisms of sound, but much more holistically, in the nature of theatre itself. For whatever reason, he nurtured this in me, and asked me to develop an introductory course in theatre for our majors. At the time, our majors took a large lecture, intro to theatre class along with non-majors, and Jim wanted to create an experience for them that was special and immersive, one that would take the impulses that had drawn them into theatre in the first place, and turn on the light bulb, so to speak. We wanted to awaken them to the magic of theatre—not just to the magic, but to the critical importance of exploring and understanding the world around us through theatre. In some ways, we wanted to do for them what now seems very much like the critical evolutionary “moment” when humans diverged from our ape ancestors—if something that took millions of years can be described as a “moment.” For our ancestors, the light bulb that got turned on was the dawning of human cognition. It caused anthropologists to assign a very specific genus to those that have such cognition to separate them from those that don’t: Homo.
I wanted to do so many things in that class, and I wanted to do them quickly. But first things first, I wanted the students to experience the joy of storytelling, something that I think is just naturally a part of the core fabric of not just theatre, but our humanity. I wanted to help them begin to think about theatre as much larger and more important than simple diversion, entertainment. At the same time, I wanted to help them develop a social bond with each other, similar to the ones I knew they had developed in high school, one that would unite them as a tribe on our campus, that would protect them from the inevitable attacks of their older peers, their non-theatre loving campus peers, their teachers and professors, even me. I wanted to help them form a group identity that would ensure their survival. In the syllabus, I wrote:
Coming to a large University for the first time and undertaking a drastic change in lifestyle, learning and commitment is a process filled with conflicting emotions—excitement/nervousness, freedom/responsibility, curiosity/insecurity, etc. Choosing a theatre major tends to compound the challenge of this experience because the commitment of the artist to the understanding of these human experiences is so great. As [Purdue professor] Rich Rand puts it, “Artists dwell on the same island as everyone else, but live closer to the volcano. They sometimes pay a mortal price for the discoveries they have made.” This course exists to help you confront these challenges.
After you have completed this course, you should recognize the significance of your relationship to Purdue Theatre. You should develop bonds with your classmates that will endure forever. You will begin to understand your own unique relationship to theatre and why it is essential for you to give theatre and life your most whole-hearted effort. You will continue down the path of life-long learning and experience, and, hopefully, learn to enjoy the many twists, forks and crossroads that make the journey worthwhile.
(Thomas 1990)
I had done a lot of work with our local parks and recreation department, and I contacted an old friend there and arranged for us to go out to one of the parks, well past closing and into the dead of night. There we built a campfire, and I asked each student to prepare and tell a story around it. We did all the ritual things one does at camp fires: cooked s’mores, told our stories—some even acted them out, reminisced about what it was that drew us into theatre, and shared who we were in a much more intimate setting than a college classroom. In short, we bonded, and created connections with each other that let each other know that they were safe in our group. An attack on one would be perceived as an attack on the many. This experience would change our relationship to each other when we returned to the classroom. It was only years later that I would realize that I had subconsciously attempted to simulate the social experience our Homo ancestors must have first had a couple of million years earlier, and probably for pretty much the same reasons.
Social experiences play a huge role in the evolution of humans. Language, music and theatre all help to develop group cooperation, coordination and cohesion, even in their most primitive manifestations. Neuroanthropologist Steven Brown argues that “the straightforward evolutionary implication is that human musical capacity evolved because groups of musical hominids out-survived groups of nonmusical hominids due to a host of factors related to group-level cooperation and coordination” (Brown 2000, 296–297). Mithen furthers the arguments Thaut made in Chapter 4 about the unique simultaneity of music by applying it to advances in social development. In Mithen’s opinion, being able to perform multiple elements of music simultaneously enables group coordination—as does being able to synchronize them to a beat, a phenomenon we’ll explore more in this chapter (Mithen 2006, vii, 115). We saw the early vestiges of these in the ape and monkey calls of Chapter 4 that were also primarily social. They contained proto-elements of language, music and theatre in signaling, vocalization/physicalization, and imitation, respectively. We survived because we developed an even more sophisticated ability to share experiences, and learn from them. We should expect to see these abilities evolve and flourish in our species as our story unfolds, then, and we do.
In this chapter, we’ll explore the evolution of our species in the context of the social pressures facing early Homo. We’ll consider how those proto-elements of language, music and theatre—signs/symbols, vocalization/physicalization and mimesis—continued to evolve. We’ll start with early Homo, and the evolution of an area of the brain known as Broca’s area that activates for both language and physical gesture. We’ll then consider Homo erectus, a much more sophisticated ancestor implicated in the harnessing of fire, and consider how this development may have led to an early lexicon and an expanding mimetic ability. We’ll consider how the evolving ability of Homo erectus to run—not walk—helped them to evolve a very sophisticated timing system useful in developing rhythm in music. We’ll consider how the development of that timing system may have given us an ability to manipulate the perception of time by others using the phenomenon of entrainment. We’ll provide some examples of how we typically use pulse and entrainment in creating sound scores for theatre. Finally, we’ll speculate on how these proto-elements of color, rhythm, mass, line and texture, words and gestures, and mimesis must have combined together to produce prehistoric rituals that would bind music and communication together in an emerging art form we would later call theatre.
One Giant Leap for Mankind
Early Homo
The earliest Homo species, Homo habilis, appeared about 2.4 million years ago in eastern and southern Africa (Smithsonian Museum of Natural History 2016).
Originally nicknamed “handyman” because of evidence found for their use of tools, later excavations found tool usage well before the known existence of Homo habilis. There have been a number of controversies over the years as to whether habilis should be categorized with Homo or with Australopithecus since they were so similar (Schwartz and Tattersall 2015). This matter has still not been resolved today, but fortunately, does not have much to do with the nature of theatre as a type of music. It simply reminds us that evolution is slow, unfailingly intertwined, and somewhat continuous.
Of greater interest to us is a related species, Homo rudolfensis, which existed about the same time as Homo habilis, but had a much larger cranium size, about 700 cubic centimeters, compared with about 500–600 cubic centimeters for Homo habilis.



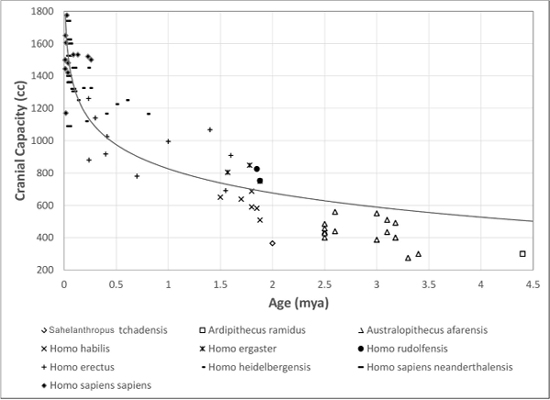
Figure 6.4 Selected Hominin cranial capacity.
Credit: Data from Schoenemann, P. Thomas. 2013. “Hominid Brain Evolution.” In A Companion to Paleoanthropology, 142–151, edited by Begun, David R. Chichester. West Sussex: Wiley Blackwell. Chart by Richard K. Thomas.
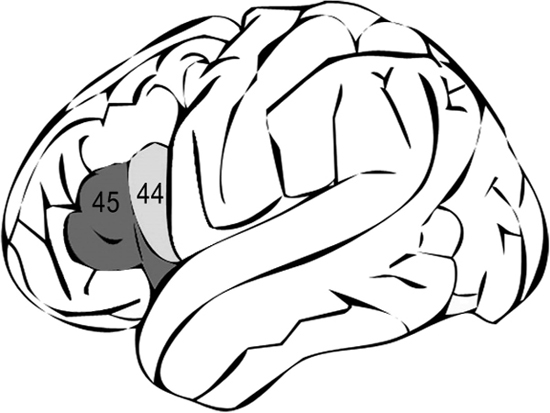
Credit: Fatemeh Geranmayeh, Sonia L.E. Brownsett, Richard J.S. Wise. 2014. “Task-Induced Brain Activity in Aphasic Stroke Patients: What Is Driving Recovery?” Brain 137 (10): 2632–2648. doi:10.1093/brain/awu163. CC-BY-3.0. https://creativecommons.org/licenses/by/3.0/deed.en. Adapted by Richard K. Thomas.

Neuroanthropologist Dean Falk examined one particular Homo rudolfensis fossil, and spotted what appeared to be evidence of an area known as “Broca’s area,” near our left temple, which activates in both speech and movement.
There are plenty of doubters on this view too, but evidence of a homologous area to Broca’s have already been found in chimpanzees, and appear to clearly show up in the next Homo evolution, Homo ergaster.
Whether or not Broca’s area clearly developed in Homo rudolfensis, or even whether Homo habilis belongs to the Homo genus, does not make such a difference for us. What makes a difference to us is that we see in these fossil remains some continuity between our cousins, chimpanzees, and the beginnings of some differences that suggest an evolving capacity for social communication.
Homo erectus
Homo ergaster roamed Africa about 1.9 million years ago, and provides us with evidence of another major leap forward in human evolution. They were tall and slender, perhaps relatively hairless, which would help improve body cooling through sweating, and had barrel-shaped chests that would provide additional muscular control over vocalizations. They also had a more human-like nose that projected outward compared to the flat nose of their predecessors. This nose provided additional resonant possibilities that could help produce a wider array of consonants and other musical expressions (Australian Museum 2015). The earliest specimen, Turkana Boy, shows a Broca’s area more clearly, possibly indicating a more advanced speech capability, better gesturing abilities, or both (Holloway, Broadfield and Yuan 2004, 139).
How language evolved during this period is also a matter of some controversy, but the divergent views all show continuity to our central thesis. Derek Bickerton argues that language consisted of two stages, the first occurring during Homo erectus, and largely consisted of just words, a developing lexicon, which we have seen somewhat characterized in the old Tarzan movies as “me Tarzan, you Jane.” He considers full mimesis to be a “spin-off” of language development (Donald 1993b, 749–750). At the same time, Bickerton recognizes that language itself must have begun as “a free-for-all, catch-as-catch-can mode that utilized sounds, signs, pantomime, and any other available mechanism that would carry intention and meaning” (Bickerton 2005, 514). Merlin Donald agrees that there may have been a “protolinguistic” stage of essentially “grammarless” speech, but argues that it would have come much later after an earlier mimetic stage (Donald 1993a, 157). Both agree, of course, that the pressure for social interaction drove the development of language.
There also seems to be little disagreement that humans would have used onomatopoeia, that is, imitations of naturally occurring sounds, to refer back to the origin of a sound source. Aarre Laakso uses the example of “grrr” as a vocalization one might use to invoke an angry dog: a vocalization that is part proto-musical (emotional) and part proto-language (referential). Laakso argues that once the use of this sound became prominent in a group, it might start to lose its immediate referent, the dog, and start to become a common symbol, for example, for anger, taking on a life of its own (Donald 1993b, 765). Mithen was so confident that some forms of imitation of animal sounds became the basis for early words and ideas that he proposed expanding “Hmmmm” to “Hmmmmm”—Holistic, manipulative, multimodal, musical and mimetic (Mithen 2006, 169–172).
Some paleoanthropologists include Homo ergaster as an early specimen in the species Homo erectus. Homo erectus lasted almost 2 million years, outliving our own species by nine times.
Homo ergaster migrated out of Africa into Asia, and possibly Europe. They had body shapes quite like ours, a much larger brain up to 800 cubic centimeters (Smithsonian Museum of Natural History 2016), and appear to be the first Hominin to use fire. Bickerton suggests that human lexicon development would have to have been in place in order to deal with the nuances of talking about fire. Bickerton also argues that in order to use fire productively, humans would have to have been able to refer to it symbolically, that is, use a word for it uncoupled from the emotion of a signaling call such as would be the case in the vervet monkey alarms1 (Bickerton, Language and Species 1990, 140–141).
Homo erectus also distinguished itself by manufacturing more sophisticated tools such as hand axes and cleavers, known as the Acheulean stone tool industry.
Arbib, one of the discovers of mirror neurons, argues that the mimesis that began with Australopithecines must have seen a period of transition from simple imitation with limited grammar to a complex imitation that included the “conscious use of pantomime with a reliance on increasingly rich memory structures capable of holding hierarchical plans for both praxis and communication” (Arbib 2013, 130–131). Merlin Donald also suggests that the development of mimetic skills in Homo erectus would have affected many areas of their social life, including “group bonding and loyalty, a much-increased capacity for social coordination, which was probably necessary for a culture capable of moving a seasonal base camp or pursuing a long hunt” (Donald 1993b, 741).


Homo erectus (and Homo ergaster) were the first species that habitually walked upright, and the first that could run long distances. It is possible that Homo erectus were able to hunt in groups because they had acquired the ability to run long distances and work in social groups using coordinated vocalizations. By working in groups, they could tire and eventually kill their prey, although this theory also has its critics (Kaplan et al. 2000, 178–181; O’Connell et al. 2002). What appears to be clear regardless of whether humans were running after prey or running for other reasons, is that Homo erectus had achieved full bipedal form, characteristically stood upright, and, unlike their Australopithecine peers and ancestors, were capable of running long distances (Bramble and Lieberman 2004). This ability would have tremendous implications for the evolution of music.
Running, Tempo, Pulse, Tactus and Entrainment
Matz Larsson is one researcher who hypothesizes that bipedal locomotion has a lot to do with the evolution of our rhythmic and musical abilities. He proposes that a main difference between early humans such as Homo erectus and our ape ancestors lies in their more modern human gait, which is characteristically more pendulum-like, a feature much closer to the steady beat required in rhythm. This gait included oscillatory movement of not just the legs, but also the arms and the head, and is typically about 120 steps a minute when walking and as much as 170 steps per minute when running. Larsson pays special attention to the sound we make when walking, having measured the sound of walking on sand and gravel as about 38 to 62 dB above the threshold of hearing. Walking or running also sends a shock wave up the body that arrives at the brain about 10 milliseconds later, a sound you can easily verify for yourself by walking or running with earplugs inserted. Walking and running are also coupled to breathing rate in all vertebrates, and you may have noticed that you run best when your breathing is synchronized in what is called a phase-locked pattern, for example breathing in on step 1, breathing out on steps 2, 3 and 4. Other patterns runners use are 3:1, 2:1, 5:2 and 3:2, with 2:1 considered the most common (Larsson 2013, 2–5). Furthermore, Nomura, Takei, and Yanagida, a group of researchers from Japan, have demonstrated that when our heart beats close to our locomotor rhythm (running steps), our heart beat will entrain to the running steps (Nomura, Takei and Yanagida 1998).
I distinctly remember the first time I ever encountered this characteristic of pacing in running. I was student teaching at East Lansing High School in Lansing, Michigan, and tasked with teaching the John Knowles classic novel, A Separate Peace, to 11th graders. To be honest, I don’t remember much about the story except that the main character, Gene, “jostled” the limb and sent his best friend Finny falling to the river bank below with a thud. But one other thing stuck out and has stayed with me all these years. Later in the book, Gene, who like me, was not much of an athlete (as compared to Finny or my brothers), was jogging around a course Finny had set up, when he suddenly, and for the first time in his life, stopped feeling the exhaustive pain of running, and suddenly felt “magnificent.” After that, running was easy. Finny explained it to Gene quite simply: “You found your rhythm” (Knowles 1960, 111–112). Quite possibly, Gene had found a pace to which both his respiratory and heart rates could entrain, a resonant frequency that simply made running easier.
This happened to me in college, and served as a turning point in my relationship with athletic activity, which I have pretty steadfastly pursued ever since in one sport or another. Jogging is a particularly interesting pastime for me, as I have been immensely fascinated about what happens when I jog and listen to music. The first thing I notice is that even if I’m not paying attention, my steps will find a way to synchronize with the beat, the basic externally provided unit of time in the music. If the tempo, that is, the speed at which the external beats of the music passes, is close, somewhere between 130 and 150 beats per minute, I’ll naturally gravitate to a one-step-per-beat rhythm. I naturally breathe in on one, and breathe out on 2, 3, and 4 during running, and in on 1 and 2, and out on 3 and 4 during heavy running.
Even more amazing, I will unconsciously find another pattern if the tempo of the music is not 130–150 beats per minute. Here is where the fabulous hemiola comes in handy. A hemiola is a musical phrase in which one rhythm is played on top of another. This could be as simple as three notes in the space of two beats, or two notes in the space of three beats. But it can also mean forcing something like two measures of triple meter against three measures of duple meter (OnMusic Dictionary 2013). When running while listening to slower tempo songs, for example in the 90–100 beats per minute range, I’ll often take six steps in the time of four slower beats of music, i.e., two measures of triple steps against three measures of two beats. In this way, every first and fourth step will always land on a downbeat (or if the meter is 4/4, on beats 1 and 3). At 110–120 beats per minute, I’ll often synchronize five beats in the space of four steps, and at a slow 80 beats per minute (or a very fast 150–160 beats per minute), I’ll even take seven steps per measure. What seems to be subconsciously important is that I land on the downbeat of each measure of the music I’m hearing. But my body really doesn’t like it when it can’t find a stepping/breathing pattern related to the music I’m listening to, and I’ll tire more quickly—a more common problem in the tempos that are between my natural running pace. Better to not listen to music than to listen to music that doesn’t sync well with my running! Fortunately, just about everything I listen to will find a way to synchronize to my running subconsciously, as I also have great flexibility in the size of my stride and how many steps per minute I take. For example, I can take short, quick steps, or long, big steps, or any combination in between, and still maintain a “running” tempo.2 And all of this applies pretty much the same to walking while listening to music, which I’ll also often do. And right on cue, we now find music streaming applications available that will find music to play that synchronize to your running tempo.
I, like so many others, find when I walk or jog to music, I’m suddenly energized, and typically lose any sense of exhaustion. Like Gene, I feel like I could just run forever. Once I’m free of the pain and exhaustion of running, my mind is free to wander to any of a number of more referential explorations, which often leads to unexpected problem solving, dreamlike mental wandering, and emotional shifts, typically toward a very simple but undeniable sense of joy. Kind of like my dreams, but now in a situation where I can consciously create the condition using music. It turns out that there is a physiological connection between my music and my running, and it’s called entrainment.
So far, we’ve focused on the process of making beats in the drumming of our hominid ancestors; we’ve not spent much time at all talking about how we experience beats. We introduced the concept of entrainment in Chapter 2, when we talked about how crabs, ants, frogs, crickets and fireflies all use it, and in Chapter 5 where we discussed entrainment in other mammals like Ronan, the sea lion, and, in our own lineage, in bonobos. This does present a bit of an evolutionary problem: entrainment seems to pop up across the animal kingdom, and yet not all that often. We must presume, then, that the fundamental characteristics of it are either hundreds of millions of years old, or happened accidentally along the way for each creature. But the type of entrainment found among the varying phyla of our distant relatives differs considerably from the human kind, because the human version does not always involve sound, has the flexibility of synchronizing not just to the beat, but to multiples and fractions of the beat, and can withstand the nuances of beats moved off their precise location, as happens in syncopation, and tempo accelerandos, and ritards (Patel et al. 2005). Tempo in music is itself rarely stable and fluctuates quite a bit compared to felt pulse or tactus, the names we give to the psychological perception of the periodic succession of beats (Thaut 2005, 9; Thompson 2014, 1097–1099). Quite frankly, human entrainment is just more sophisticated.
As I’m now writing this chapter, I’ve been especially conscious of examples of entrainment occurring in my day-to-day waking world. A couple of weekends ago, I went to a local jazz concert in a small venue in town where a lot of local musicians also gathered. As we watched the musicians play, and listened to their music, I started looking around for signs of entrainment, the biological process in which our motor reactions are modified by exposure to external rhythms. The signs were everywhere. Surprisingly, the signs of entrainment manifested themselves in myriad ways. Just about everyone moved to the beat, but they didn’t all move in exactly the same way. A significant number of people simply tapped their feet to the music. Some tapped their toes to the primary pulse, others doubled down and tapped twice to each pulse. A few only tapped once every other beat. One of my musician friends tapped on the first, third and fourth beat of each measure, creating her own “silent” contribution to the music. Then there were the head boppers. Some nodded front to back, some side to side. They were augmented by the finger tappers. Some tapped on their tables, some on their laps, some flammed their taps from baby finger to index finger. Some combined these; some got their whole body involved in a sort of seated dance. The effect of the band’s music on the audience was multimodal. The auditory beats somehow found a way to create corresponding motor reactions in a wide variety of ways in the audience. It would have been interesting to shut off the music and close-mike every single person’s movement and play it back together. Why? Because it would have all been pretty much in sync musically.
What’s even more astounding about this phenomenon is that most of the audience members were unlikely to have been aware that they were physically moving to the beat. I know this because I’ve played music in my classes, and watched students react. Then I stopped the music and specifically asked them if they were aware of their physical movements. Some were, but the vast majority didn’t even realize they were doing it. That’s entrainment, and it is one of the most important techniques we use to manipulate a person’s perception of time and, together with other elements of music, their emotions and moods.
Neuroscientist Michael Thaut discovered that our biological reaction to external rhythms is not what we always thought it was. We used to model rhythm and rhythmic formations as clocks, pulse counters or stopwatches. Now we understand that pulses from external music fundamentally entrain oscillatory circuits in our brains. Our brain encodes the neural firing rates of the auditory system and projects those rates on to other resonant brain tissue (Thaut 2005, 6–7). In other words, an external audible pulse is the tempo of our music; it modifies key circuits in our brains to oscillate at the rate of the external tempo. We don’t simply imitate the external auditory pulses, we become them. That same direct connection we talked about in our primitive brain limbic system and cerebellum that made us jump at a startle event in Chapter 2, allows external rhythms to unconsciously manipulate us physically. We change. And it happens subconsciously—Harold Burris-Meyer’s “You can shut your eyes, but the sound comes out to get you” is at it again! You can fight it with your conscious frontal lobe, but the moment you stop, it will creep back in again. Without even thinking, we start moving rhythmically in response to the external pulses of the music. That’s different than imitation, where we make a conscious effort to say “alright, I go like this, and you do the same.” This is entrainment, and it’s a major function of music, and one that distinguishes music from the referential functions of language and images. We don’t tap our feet to conversation. And it seems to be an area in which our auditory systems excel over our visual systems in perception of time; it’s much harder for us to consciously synchronize to a visual pulse than an auditory one (Patel et al. 2005, 226; Thaut 2015, 256).
We as humans have evolved quite sophisticated abilities to create, manipulate and synchronize to beats. Our most fundamental sense of this is called pulse or tactus, the level of beats to which entrained humans typically tap their feet (Roholt 2014, 86). Tempo, then, refers to how the music is played in beats per minute, whereas tactus refers to how music is experienced. Tactus is an interval, period-based process expressed as the duration of each pulse (Thaut 2005, 8). For example, if we tap our feet 120 times per minute, then we would perceive two pulses per second, one every half a second, a 500 millisecond pulse. Humans seem to be able to perceive pulse intervals between 200 and 2,000 msec (30–300 beats per minute) (Grahn and Brett 2007, 894). However, humans tend to perceive tempos outside the region of 60–150 bpm as multiples or subdivisions of tempos in the 60–150 bpm range (Thaut 2005, 9).
Not all music is based on beats, of course. For example, a lot of ambient music consists of long flowing tones without a well-defined beat. We call this unmeasured music, as opposed to music with a well-defined tactus, which we call measured music. Measured music can be further subdivided into steady pulse and mutable pulse varieties. Most folk, dance and pop music typically has a steady pulse, whereas classical music often has a very changeable pulse that includes rubatos, ritards and accelerandos (Merker, Madison and Eckerdal 2009, 8).
Our perception of pulse, and how we react to it is fairly complicated. To begin with, it makes a difference whether we are consciously attempting to synchronize with a beat; for example, if we are attempting to play a piece of music along with someone, rather than simply listening to and enjoying music.
Simply listening to music activates brain circuits that automatically entrain to pulses. This is the part where we start tapping our toes without ever consciously thinking about hearing the beat. These automatic timing circuits are the ones we often try to tap into as playmakers; the ones where we can affect our audience without them having to pay attention to what we are doing musically. In scoring, we often want our audience to concentrate on referential images and language, and not to pay any attention at all to the music. The auditory system provides input to what are called the motor areas of the brain in the frontal and parietal lobes, roughly at the top center of our brain.
Without much help from our conscious brain, neural networks within a broad area of structures that include these motor centers, the spinal cord, the basal ganglia, and the cerebellum, match the timing of the audible pulses and send them down our spine to the various parts of our body physically creating the same beats, for example, by tapping our feet (Thaut 2015, 257). Researchers expect that our sensorimotor system (combination of auditory and motor systems) gets involved by simply copying the interval between pulses and then repeating them. However, this process only seems to work with small differences, temporal errors much less than the amount of time between the two most recent pulses, a process called phase correction3 (Repp and Keller 2004, 500). Generally speaking, as long as the tempo doesn’t change too drastically, our motor circuits correct our motor reactions to coincide to whatever the length of time was between the last two beats. We are constantly adjusting to the beat without even knowing it! Of course, we can cognitively control this also, but generally speaking, most humans can move to a tempo without consciously even thinking about it.
If things start to get too far off, or we decide we want to participate in a different way, such as tapping our hand instead of tapping our foot, our cognitively controlled timing circuits get involved in a process called period correction, which may average the duration of multiple pulses to find the new tactus. These circuits require conscious involvement from other parts of our frontal and parietal lobes associated with our memory and attention. Playing music and synchronizing our beat to another musician requires the use of both automatic timing circuits and cognitively controlled timing circuits. In practice researchers imagine that there is a “close and frequent” interaction between automatic and cognitively controlled timing systems (Merker, Madison and Eckerdal 2009, 11; Repp and Keller 2004, 501).
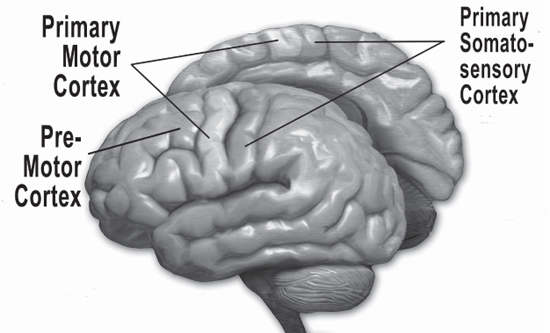
Credit: “http://Blausen.com” Blausen.com staff 2014. “Medical gallery of Blausen Medical 2014”. WikiJournal of Medicine 1 (2). doi:10.15347/wjm/2014.010. ISSN 2002–4436. CC-BY-3.0 (https://creativecommons.org/licenses/by/3.0/deed.en). Adapted by Richard K. Thomas.
Tempo, Pacing, Tactus, Entrainment and Theatre Composition
When we create underscoring, we want the audience to focus on the referential language, images and action of the scene rather than on the music. However, when there’s a show-stopping “song and dance” number in a musical, or even a simple song in a Shakespearean play, we will often want the audience to focus on the musical aspects of the play as well. So, it helps to know when we want to stimulate our automatic timing circuits (for example, by using a steady tempo), and when we want to draw conscious attention to the music (for example, by inserting substantial tempo changes). It does help to know that phase correction using our automatic timing circuits is very hard to suppress, which means we habituate to it quickly, and, as long as we don’t start monkeying with the tempo too much, it will quickly return to its automatic, unconscious function (Repp and Keller 2004, 517–518).
One of the most common requirements in which sound typically carries the music of a theatre piece all by itself is scene change music. The term “scene change” seems to have been colloquially developed over a long period of time, but in actuality, the term can often be a misnomer, as the scene—the place where an incident occurs (Oxford Dictionaries 2016)—sometimes doesn’t really change at all, for example, in a single unit set such as a drawing room comedy. What must change in virtually every case is time, and we use music to control our perception of time. So, in practice, I like to think of scene changes as tactus changes.
At the most fundamental and basic level, we are charged with changing the underlying pulse of the departing scene to that of the entering scene. To understand the difficulty in accomplishing this, we must first understand that the tactus of a given scene is quite mysteriously created by a combination of contributing musics.
First, there is the implied tactus created by the prosody of the actor’s voices; the speech rhythms, melodies, dynamics and so forth that actors have carefully created over a long period of rehearsing, often referred to in theatre as pacing. Experienced sound designers, who spend a lot of time in rehearsals, know to expect certain changes in pacing as rehearsals progress into performances. At the first read-through, the pacing is often much faster than it will wind up being in performance. This is, of course, caused by the fact that the actors have not integrated the prosody of their speech with the music of their physical movements. Once they get in rehearsal, the actors start moving in time, creating a visual dance, that, for a while, slows down the pace—usually a bit too far, as the actors learn the distinct rhythms. This is not unlike how a piano player learning how to play a complicated section of music will first slow down to a speed they can play without mistakes. As the actors figure out their rhythms, their pacing will pick back up again until tech rehearsal, when the physical process of moving in and adapting to a new space will slow things back down again. Finally, during previews, the actors will tend to fully internalize the mimesis of the performance and the pace will stabilize, although the audience and other personal factors will still cause slight variances around the more stable pacing of the show. I remember the stage manager of Tommy on Broadway boasting that the pacing of the show was so stable (i.e., the music was performed to a “click” track) that the running time of each performance was within 30 seconds every evening! This is not typically the case, however, and experienced sound designers use specialized software and know-how to build cues and music to allow their sound scores to breathe musically within the slightly varying tempos of each performance.
Then there is the tactus created by the world inhabited by the actors; a world created by set, costume and light designers that may or may not correlate to the tactus implied by the script, and rehearsed by the actors. Phillip Silver eloquently addressed this issue in his 2003 address to the World Congress of OISTAT, in which he described the moving of a comedy from a theatre with a 9.5 meter proscenium to a 12.2 meter proscenium, a seemingly small change that required the actors to stretch out their physical timing—rhythms—to the larger space. That stretching of rhythms destroyed the comic timing the playwright had written into the smaller proscenium show (Silver 2003). On a more positive note, I designed sound for a production of Athol Fugard’s The Island that played in a number of regional theatres for a couple of years, in which director/set designer Jim O’Connor constrained the set to a 10 foot by 10 foot square, thus forcing the actors’ physical and vocal rhythms into the cramped confines needed to fully realize the abject despair of their imprisonment. The impact on the tactus of the actors was positively chilling, forcing short, chopped rhythms in a space with no room to move.
Once sound designers and composers understand the nature of all of the various rhythms, pacings and tempos interacting in a scene, they are in a good position to create “scene (or tactus!) change” music. First, they must have a fairly good sense of the tactus of both the composite outgoing and incoming scenes. Oftentimes they must deduce or predict this from just the printed words on the pages of a script. If they are lucky, they will get to participate in the rehearsal process themselves, and be able to develop firsthand a strong sense of the required tactus for the scene change.4 More frequently, however, they must be prepared to “adapt” that tactus on the fly, especially when the tactus dictated by the arrival of a physical set and costumes during tech rehearsals also begins to integrate with the pacing developed by the actors.
To accomplish the tactus change, one of our most common techniques is to introduce a bit of music under the end of one scene, typically at the approximate tactus of the scene. If we nail the tactus correctly, then our introduction of music will habituate quickly, and the emotional quality of the music will incite a similar mood in our audience. At the scene change, then, we might abruptly change the tactus, drawing our audience’s attention to it, which, of course, causes them to stop focusing on the outgoing scene. We establish a new tactus during the scene change, perhaps inducing a little bit of entrainment in the process, and then we finally introduce a new tactus for the incoming scene, hopefully creating a sense of the pulse in the audience appropriate for the actors’ pacing at the commencement of the new scene.
But not always. There are perhaps an infinite number of variations on this process; if there weren’t, the Tony Awards Committee would have been justified considering that sound design was not an art form worth including back in 2014.5 Sometimes the creative team will want to introduce the tactus of the incoming scene during the outgoing scene, to begin to allow the rest of the music to impinge its emotional stimulation on the outgoing scene. Sometimes the team will do just the opposite: hold the tactus from the outgoing scene through the scene change and into the new scene, refusing to let go of the emotional qualities of the prior scene even as the incoming scene starts. And, of course, some times the team will introduce a thematic tactus, a pulse that they have used elsewhere in the play to subtly incite an audience recollection of a prior scene or event while still immersed in the current scene (more on this later, when we take up the subject of music’s close relationship to memory). And, finally, they may choose no audible musical transition at all, preferring to allow the existing tactus of the outgoing scene to transition on its own to that of the incoming scene.
To multiply the possible options, consider that the creative team may make a conscious choice to contrast the current tactus of a scene with quite a different tactus. For example, a scene that ends in a major fight might be underscored with a piece of music with a very slow tempo, or a fight to the scene-ending death might be staged in slow motion while the music plays at a very fast tempo. The ability to contrast the referential content of a scene with the emotions and sense of time incited by the music creates an almost unlimited array of possibilities that can only be truly brought to life by a team that has the ability to imagine the possibilities.
In addition to understanding and designing around the existing pacing and tactus of outgoing and incoming scenes, we also need to manipulate our audience’s perception of time, so that we become aware that a certain amount of time has passed between the outgoing and incoming scenes. To accomplish this, we turn to the more conscious method of entrainment employed in pulse manipulation, and period correction, in which we employ tactus changes large enough to attract our audience’s attention. In its most simple form, we introduce a piece of music at the scene change that draws attention to itself. The audience soon entrains to the new music, forgetting what just happened in the outgoing scene. We end the scene change music and the next scene starts, establishing its own tactus. The duration of the scene change music, coupled with the shift in tactus that possibly includes its own entrainment, instills in the audience the sense that a certain amount of time has passed.
A long time ago producing radio dramas, I learned an old trick: how long the scene change music lasts helps the audience understand how much dramatic time passes between scenes. It’s a very simple rule: if the next dramatic scene happens only a few moments later, do not have any scene change time at all. If the dramatic time is a few hours long, then have a relatively short scene change time. If the dramatic time was days, weeks or months long, create progressively longer scene changes, even consider adding a voiceover to help create the sense of time passing. In short, the longer the scene change time, the longer the dramatic time will appear to have passed. It’s a simple and obvious rule, and one that complements Einstein’s thoughts about the relativity and subjectivity of time quite well.
We don’t tend to use this rule in theatre. In theatre, scene changes are largely determined by the decidedly undramatic amount of time it takes to change either the scenery or the costumes. It’s somewhat understandable to have to extend the dramatically useful amount of scene change music if the scene change can’t be performed in a short amount of time, but it has always been a bit baffling why we would shorten the scene change time if the scene change would not take as long as required dramatically.
A few years ago, I had the great pleasure to work with Janet Allen, artistic director of the Indianapolis Repertory Theatre, on a production of Ernest Thompson’s American classic, On Golden Pond. Janet was understandably concerned because the single-set play required lengthy scene changes of both set and costumes due to the fact that each change took place at a considerably later point in dramatic time. For example, the set and costumes would have to change from spring to summer, even though the summer cottage setting did not fundamentally change. How would we fill the time? In our early conversations, Janet confided in me that she hates lengthy scene changes, because they allow the audience to disconnect from the play, and once disconnected, are very hard to get back. Instead, we talked about a way to distract the audience (my term), which is quite different from allowing them to disconnect. For our scene changes, we would use Carrie Newcomer music. Newcomer, a Grammy award-winning singer/songwriter, had been a long-time collaborator of mine (which I’m sure had a lot to do with why Janet hired me to design the sound in the first place!). Carrie writes provocative lyrics that poignantly resonate in themes such as those underlying On Golden Pond. Our strategy would be to distract the audience, getting them to focus on the thoughts and inspirations in Carrie’s music, while the scene change unfolded in front of them visually for a bit of added distraction. There’s nothing earth-shaking about this technique, but curiously, I had designed sound for close to 40 years without ever having a conversation with a director about how the technique actually works. In this case, the really important reason the technique works is because dramatic months pass in between the scenes, and providing these breaks helps the audience leave a scene and enter a new scene with a subjective sense that much more time has passed.
But at a subtler level, the scene change music itself created a rhythmic entrainment in the audience—it was literally long enough that you could see the physical evidence in the audience unconsciously moving in time to the music—the new tactus introduced to grab control of the audience’s perception of time. Choosing the distracting music was a challenge, but not the hardest part. The most challenging part was figuring out how to blend the tactus of the departing scene into the distracting music, and then how to let go of the tactus of the distracting music in order to establish the tactus of the incoming scene. For example, in one scene we transitioned out of a moderate tempo Carrie Newcomer piece into an ambient tactus created by bullfrogs—themselves entrained at a very slow tempo—in order to bring the tactus of the incoming scene down to a much lower energy level, which in turn gave us a place to build a new rising action (increasing pulse) for the following scene. Embedded in all of this was a great need to create in the audience this sense of a substantial amount of time passing, but without letting them disconnect from the play in a way that would be very hard to get them back.
In tactus, and by extension to its physicalizing counterpart, entrainment, we witness another clue to the flexibility of time itself. Michael Thaut tells us that “Pulse rate defines, within given interval durations, how fast time, expressed through music, flows…. The primary element in music that creates the perception of time is rhythm” (Thaut 2005, 9, 15). Rhythm liberates us from the tyranny of a chronological clock; our brains and bodies are really not concerned with that in theatre anyway. While circadian rhythms, our biological entrainment to the cycle of night and day, have been a part of our core existence since the very earliest organisms, the division of that cycle into somewhat arbitrary periods of hours, minutes and seconds has little to do with our biological timing mechanisms (Scientific American 2007). Quite the contrary: in theatre, we may experience the passing of 24 hours, or even days, months or years, within the short chronological time of a couple of hours, just like many people report they do in dreams.
Entrainment suggests that our unique and subjective perception of time is more related to a diverse number of oscillatory circuits in our brain, especially the ones exterior pulses entrain to that cause motor movements in our bodies. Consider that we tap our foot the same number of times in 60 seconds at 60 bpm as we do in 30 seconds at 120 bpm. We are quite aware that it takes more time to tap 60 times in 60 seconds than it does to tap 60 times in 30 seconds. And yet, the same number of events take place in either case! Our perception of time is relative; time passes slower at slow tempos and faster with fast tempos, rather than being constant. Sometimes that’s a good thing, and sometimes, not so good. Who among us has not witnessed a rehearsal in which the director begged the actors to “pick up the tempo”? Why? Because, at least for the director, and presumably for the audience, time seems to be passing incredibly slowly. Conversely, when the tempo is too fast for a scene, we disengage in a similar way: we don’t fully transport into the world of the play; time is simply passing too fast for us to fully engage. But when the actors and musicians hit what I’ll call the “resonant” tempo of a scene, that is, the ideal tempo, something magic happens: we no longer think about the passage of time, and enter that dreamlike state in which the world outside of the of the play ceases to exist and we are fully transported to the dramatic time and space of the performance.
We use tactus to transport our audience there in time. We use tactus to manipulate our audience’s perception of time, knowing that our perception and our experience of time is unique to each individual, but ever determined to bring those individual perceptions more into a common one. Playwrights, directors, actors, designers—both visual and auditory—and composers all use tempo, pacing, pulse, tactus and entrainment, both knowingly and unknowingly, wittingly and unwittingly, to coax a common perception of time out of our audience. Since time is relative, we are keenly aware of how we are manipulating the pulse of our plays around what came before, and what comes next, for example, as part of the “rising action” of traditional linear plays. As we will see in later chapters, manipulating tactus is one of the ways in which we maintain arousal in our audience and hold their attention.
Our brains are keenly able to measure—anticipate—time via the individual length of pulses in external tempo/pacing and internal tactus. They physically respond to such pulses. When we develop the tactus of a play, we are working to pull our audience toward a common perception of time, oblivious to the ticking of the chronological clock outside of our dramatic experience. Rhythm is one of the most powerful tools we use to transport the audience from the external world of lobbies and house lights and our workaday problems to the more dreamlike world of the play. Referential visual images and language compel our audience to “suspend their disbelief,” but participation in the tactus of the scene, which is almost always accompanied by a strong emotional reaction incited by the music of the play, carries those images and ideas into the heart and soul: Music as a Chariot.
When Music Meets Mimesis
Homo erectus migrated over a fairly large part of the world for almost 2 million years before finally dying out about 143,000 years ago. In order to migrate successfully, they must have developed tremendously in terms of their music, language and ritual abilities, and would have had to develop tremendous social abilities to hold their tribes together against the extraordinary pressures of both nature and predators. Imagine then, how all of this may have come together around a campfire somewhere in that distant past, a campfire not unlike the one we built to help my new theatre majors bond in our not so distant past….
Presumably the tribe would gather around that campfire to cook the food they had gathered, hunted, scavenged or foraged. And while hanging out around that campfire, what do we imagine they did, just sit around and grunt? That hardly makes for a pleasant evening of entertainment; one imagines a much more socially active scenario. Daniel Levitin suggests that “singing around the ancient campfire might have been a way to stay awake, to ward off predators, and to develop social coordination and social cooperation within the group” (Levitin 2007, 183). Semiologist6 Jean Molino proposed a common underlying phenomenon that would create extraordinary social cohesion around a campfire: rhythm. Molino imagined “a group of hominids would perform activities of collective imitation without language but accompanied by vocalizations and organized by rhythm: these would in fact be the first forms of the representation of scenes, that is, of narratives, leading to rite and to myth” (Molino 1999, 174). A very early linguist and feminist, Jane Harrison, found a clear link between just such an activity as this and theatre:
When the men of a tribe return from a hunt, a journey, a battle, or any event that has caused them keen and pleasant emotion, they will often re-act their doings round the camp-fire at night to an attentive audience of women and young boys. The cause of this world-wide custom is no doubt in great part the desire to repeat a pleasant experience; the battle or the hunt will not be re-enacted unless it has been successful. Together with this must be reckoned a motive seldom absent from human endeavour, the desire for self-exhibition, self-enhancement. But in this re-enactment, we see at once, lies the germ of history and of commemorative ceremonial, and also, oddly enough, an impulse emotional in itself begets a process we think of as characteristically and exclusively intellectual, the process of abstraction.
(Harrison 1913, 42)
An emotional impulse begets an intellectual process? Sounds like music carrying ideas to me. Imagine a group of Homo erectus gathering around a campfire to cook the evening meal. While waiting for the meal to cook, one of the leaders of the hunt, or maybe the “class clown,” gets up and starts to imitate the hero leading the charge to take down the prey. And then something magical happens….
Evidence for musical instruments is hard to come by and won’t appear in the archaeological record for another hundred thousand years or more—maybe a lot more. But stones are easy to come by, and it wouldn’t take much for a couple of members of the tribe to start pounding a couple together, egging our protagonist on to a rhythmic beat. Bones from prior kills would make even better percussion tools, despite the fact that they didn’t have “Vic Firth” engraved on the side of them. They are hollow, more resonant, make a great sound when struck together, and raise the mystique of the hunt in a way that is not completely referential. Pretty soon everyone in the tribe is participating. Evolutionary psychologist Geoffrey Miller reminds us that music in tribal societies is almost always a group affair with everyone participating and no one simply sitting and listening (Miller 2000, 348). We can imagine everyone participating vocally too, imitating the sounds of the hunters, the sounds of the prey, perhaps they’ve even developed words and names for the prey, food, warriors and more and have learned to chant them in time to the beat. All of this emoting musically to a beautiful crescendo of socially connected cacophony.
And then it happens. The participants entrain to the beat, their bodies undulating to the external tempo of the pulsating beat. They transcend mimesis and become dancers, however simple, however primitive. They cease to be observers of the imagined hunt, and become participants. They enter the dreamlike state in which they quiet the parts of the prefrontal cortex that consciously remind them of their present surroundings. They indulge their more primitive cerebellum and limbic systems, thus creating internal mental images conjured by the imitations, and fueled by the entrainment that rhythmically and physically connects their bodies to the protagonist and the rest of the group. It is in that moment when they leave the scene around the campfire and enter the mise-en-scène7 of their minds. They are no longer audience members watching a ritual unfold. They are no longer simply imitating. The become active participants in not just the ritual, but in the imaginary hunt itself. They have transported back in time to the hunt; they do not “relive” the hunt, they simply “live” it in the moment; for them it is actually happening, their perception of time controlled by the music and the entrainment. Some elements playing out in their dreamlike state coincide directly with the perception of everyone else in the group, creating a strong and impenetrable social bonding. But other elements are unique to each individual’s experience and perspective, for example the visualization of the world in which this hunt took place. It is in this moment of music meeting mimesis that theatre is born. And music is the chariot that transports our ancient participants to this very real and yet very imaginary world.
Conclusion
In this chapter, we have traced the early evolution of our human ancestors, our genus Homo. In Homo we discovered evidence of evolving forms of music, language and theatre in the emotive expressions, in signs and an emerging lexicon, and in the mimesis of increasingly sophisticated rehearsals, re-creations and new creations; in the performances of imitative gestures and vocalizations. We paid special attention during this period to how the simultaneous evolution of the ability of humans to run—not walk—may have led to tempo, pace, pulse, tactus and human entrainment, the special ability to create a beat, and to synchronize that beat with others. We explored the unique connection between our auditory system and our motor system in entrainment, in which auditory pulses subconsciously cause physical movements in subjects. Without even thinking, we tap our toes to an external beat. Along the way, we’ve explored how these evolutionary traits manifest themselves even today in choices we make creating sound scores for theatre. In particular, we’ve explored how we use external tempos to manipulate our perception of time and pacing. Finally, we’ve hypothesized how the simultaneous evolution of music, language and mimesis may have manifested themselves around the campfire, how much more sophisticated ritual may have emerged, and how ritual bore the primal characteristics of theatre and dance, emerging art forms based in music, language and mimesis.
Ten Questions
- Describe four early forms of the genus Homo and the major evolutionary developments of each that relate to music, language and theatre.
- What is a major difference between Bickerton’s evolutionary hypothesis for language development in Homo erectus and Donald’s/Arbib’s?
- How does human entrainment differ from entrainment found in vertebrates such as frogs and fireflies?
- How does tempo differ from the internal felt pulse?
- Define rhythmic entrainment.
- How is the multimodal nature of human entrainment different from that of our vertebrate ancestors?
- How does entrainment demonstrate that our brain does not treat rhythm like a clock or a pulse counter?
- Describe the differences between beat, pulse, tactus and tempo.
- How does entrainment differ from imitation?
- What are two different kinds of entrainment, and what are their characteristics that distinguish them from one another?
Things to Share
- Download an app that allows you to tap out a beat and then tells you the tempo. Put together a playlist for a walk or run that includes music to which you think you might like to enjoy walking or jogging. Record the name of the song, artist, and beats per minute (bpm) of your playlist in the following spreadsheet. Put on a pair of closed ear headphones or earplugs. Don’t pump any music through them yet. Just go out for a walk or a jog and pay attention to the tempo with which you typically like to walk or jog. Determine your natural walking or running tempo and make a note of that. Start your playlist, and go on your walk or run. Try not to pay attention to the relationship of your steps to the beat. Instead, consciously think about something else for a while. Eventually (say halfway through the song), let your attention turn to whether your body has unconsciously found a relationship between the beat in your music and your footsteps, and what that relationship is (one beat per step, three steps per two beats, etc.). Make mental notes about music that helped you run easier, and music that seemed to make running harder. When you finish your run, go back and make notes on the spreadsheet below about how many steps per beat you found yourself taking during each song, how many steps per breath you found yourself taking and any other notes. Which songs helped your run or walk, and which made it harder? What was the relationship between your steps per minute and your beats per minute? How did your breathing play into your comfort zone?

- Find a movie that has won an Academy Award for sound editing. Pick a movie that you love and wouldn’t mind seeing again. Now get hold of the movie (perhaps buy it, because you can use it for other projects in this class, rent it from Netflix, or even check it out from your local library). Watch it with an ear toward a composer consciously using entrainment, and, if possible, manipulating the pulse in a tactus change. Make a note of the time in the movie where the entrainment or tactus change occurs so we can cue it up in class and you can share it with us. Pay careful attention to how the sound artists manipulate changes in the tempo of the sound score to manipulate the sense of time in the listener. Be prepared to describe for us how you think the sound team is manipulating us. Bring at least two examples with you in case someone uses your first example.
- Having explored the wonderful world of mass (volume, shape, dynamics), now it’s time for you to explore these concepts in composition. Compose or design a 1:00–1:30 piece that does not use a well-defined tempo, only uses one color such as a guitar or piano, and focuses on manipulation of the dynamics and volume of the moment. Find a place to introduce both a habituation and a startle moment. Play with crescendos and decrescendos. How close can you get to the noise floor of the listening environment, and how can you make the other extreme—the overloading of your meters—seem all the more loud because of how quiet your composition got? As always, play and experiment!
Notes
1It is worth noting that, as of this writing, the earliest hard evidence we have of Homo erectus using fire is only 1 million years ago, leaving a gap of almost a million years between their arrival and their use of fire (Berma et al. 2012, E1215. Also, see James 1989).
2Running being defined as a form of locomotion in which one of my feet is always in the air.
3Phase in this case simply means the difference in time between where in time the next expected pulse should start, and where it actually starts, typically measured in degrees with 0° meaning no error and 360° meaning one whole pulse off.
4If they are really lucky, they will be composing the music to a musical or opera, or other form in which the composed music dictates the tempo of the tactus change!
5Fortunately, the Tony Committee came to their senses (literally!), and as of this writing in 2017, have reinstated the Tony Awards for Sound Design.
6A semiologist is a person who studies semiotics (i.e., sign processes and meaningful communication).
7A French term that literally means “putting on the stage.” In modern theatre it typically refers to the composite visual images created and experienced in a play. Our point in this example, of course, is that the roots of theatre do not lie in the visual elements of the play; they are created in the minds of the audience by attaching ideas to music.
Bibliography
Arbib, Michael A. 2013. “Précis of How the Brain Got Language: The Mirror System Hypothesis.” Language and Cognition 5 (2-3): 107–131.
Australian Museum. 2015. “Homo Ergaster.” September 25. Accessed April 2, 2016. http://australianmuseum.net.au/homo-ergaster.
Berna, Francesco, Paul Goldberg, Liora Kolska Horwitz, James Brink, Sharon Holt, Marion Bamford, and Michael Chazan. “Microstratigraphic Evidence of in Situ Fire in the Acheulean Strata of Wonderwerk Cave, Northern Cape Province, South Africa.” Proceedings of the National Academy of Sciences of the United States of America 109 (20): E1215-20.
Bickerton, Derek. 1990. Language and Species. Chicago: University of Chicago Press.
———. 2005. “Language Evolution: A Brief Guide for Linguists.” Science Direct 117 (3): 510–526.
Bramble, Dennis M., and Daniel E. Lieberman. 2004. “Endurance Running and the Evolution of Homo.” Nature 432 (7015): 345–352.
Brown, Steven. 2000. “The ‘Musilanguage’ Model of Music Evolution.” In The Origins of Music, edited by Nils L. Wallin, Björn Merker, and Steven Brown, 271–300. Cambridge, MA: MIT Press.
Donald, Merlin. 1993a. “Human Cognitive Evolution.” Social Research 60 (1): 143–170.
———. 1993b. “Précis of Origins of the Modern Mind.” Behavioral and Brain Sciences 16 (4): 737–791.
Grahn, Jessica A., and Matthew Brett. 2007. “Rhythm and Beat Perception in Motor Areas of the Brain.” Journal of Cognitive Neuroscience 19 (5): 893–906.
Harrison, Jane. 1913. Ancient Art and Ritual. New York: Henry Holt.
Holloway, Ralph L., Douglas C. Broadfield, and Michael S. Yuan. 2004. The Human Fossil Record. Vol. 3. Hoboken: John Wiley & Sons.
James, Steven R. 1989. “Hominid Use of Fire in the Lower and Middle Pleistocene: A Review of the Evidence.” Current Anthropolgy 30 (1): 1-26.
Kaplan, Hillard, Kim Hill, Jane Lancaster, and A. Magdalena Hurtado. 2000. “A Theory of Human Life History Evolution: Diet, Intelligence, and Longevity.” Evolutionary Anthropology 9 (4): 156–185.
Knowles, John. 1960. A Separate Peace. New York: Bantam.
Larsson, Matz. 2013. “Self-Generated Sounds of Locomotion and Ventilation and the Evolution of Human Rhythmic Abilities.” Animal Cognition 17 (1): 1–14.
Levitin, Daniel J. 2007. This Is Your Brain On Music. New York: Penguin Group/Plume.
Merker, Bjorn H., Guy S. Madison, and Patricia Eckerdal. 2009. “On the Role and Origin of Isochrony in Human Rhythmic Entrainment.” Cortex 45 (1): 4–17.
Miller, Geoffrey. 2000. “Evolution of Human Music Through Sexual Selection.” In The Origins of Music, edited by Nils L. Wallin, Björn Merker, and Steven Brown, 329–359. Cambridge, MA: MIT Press.
Mithen, Steven. 2006. The Singing Neanderthals. Cambridge, MA: Harvard University Press.
Molino, Jean. 1999. “Toward an Evolutionary Theory of Music and Language.” In The Origins of Music, edited by Nils L. Wallin, Björn Merker, and Steven Brown, 165–176. Cambridge, MA: MIT Press.
Nomura, Kunihiko, Yoshiaki Takei, and Yasuyoshi Yanagida. 1998. “Analysing Entrainment of Cardiac and Locomotor Rhythms in Humans Using the Surrogate Data Technique.” IEEE Engineering in Medicine and Biology Magazine 17 (6): 54–57.
O’Connell, J.F., K. Hawkes, K.D. Lupo, and N.G. Blurton Jones. 2002. “Male Strategies and Plio-Pleistocene Archaeology.” Journal of Human Evolution 42 (6): 831–872.
OnMusic Dictionary. 2013. “Hemiola.” February 14. Accessed March 11, 2016. http://dictionary.onmusic.org/terms/1697-hemiola_36.
Oxford Dictionaries. 2016. “Scene.” Accessed April 2, 2016. www.oxforddictionaries.com/us/definition/american_english/scene.
Patel, Aniruddh D., John R. Iverson, Yanqing Chen, and Bruno H. Repp. 2005. “The Influence of Metricality and Modality on Synchronization With a Beat.” Experimental Brain Research 163 (2): 226–238.
Repp, Bruno H., and Peter H. Keller. 2004. “Adaptation to Tempo Changes in Sensorimotor Synchronization: Effects of Intention, Attention, and Awareness.” Quarterly Journal of Experimental Psychology 57 (3): 499–521.
Roholt, Tiger C. 2014. Groove: A Phenomenology of Rhythmic Nuance. New York: Bloomsbury.
Schwartz, Jeffrey H., and Ian Tattersall. 2015. “Definining the Genus Homo.” Science 349 (6251): 931–932.
Scientific American. 2007. “Why Is a Minute Divided into 60 Seconds, an Hour into 60 Minutes, Yet There Are Only 24 Hours in a Day?” March 5. Accessed March 24, 2016. www.scientificamerican.com/article/experts-time-division-days-hours-minutes/.
Silver, Phil. 2003. Communication of Space and Place—35 Years of PQ. Prague, June 13. Unpublished.
Smithsonian Museum of Natural History. 2016. “Human Evolution Timeline Interactive.” March 25. Accessed March 28, 2016. http://humanorigins.si.edu/evidence/human-evolution-timeline-interactive.
Thaut, Michael H. 2005. Rhythm, Music, and the Brain. New York: Routledge Taylor & Francis Group.
———. 2015. “The Discovery of Human Auditory-Motor Entrainment.” In Music, Neurology and Neuroscience: Evolution, the Musical Brain, Medical Conditions and Therapies, edited by Eckart Altenmüller, Stanley Finger, and Françoise Boller, 253–266. Amsterdam: Elsevier.
Thomas, Richard K. 1990. “THTR 202 Fall 1990 Syllabus.” West Lafayette, IN: Unpublished, August 16.
Thompson, William Forde, ed. 2014. Music in the Social and Behavioral Sciences. Vol. 1: Los Angeles: Sage.