
9.2.4 Batch Reactor Calculations for Enzyme Reactions
A mole balance on urea in a batch reactor gives
![]()
Because this reaction is liquid phase, V = V0, the mole balance can be put in the following form:
![]()
The rate law for urea decomposition is
![]()
Substituting Equation (9-31) into Equation (9-30) and then rearranging and integrating, we get

We can write Equation (9-32) in terms of conversion as
Curea = Curea0 (1 = X)

The parameters KM and Vmax can readily be determined from batch reactor data by using the integral method of analysis. Dividing both sides of Equation (9-32) by (tKM/Vmax) and rearranging yields
![]()
We see that KM and Vmax can be determined from the slope and intercept of a plot of (1/t) ln[1/(1 = X)] versus X/t. We could also express the Michaelis–Menten equation in terms of the substrate concentration S:

where S0 is the initial concentration of substrate. In cases similar to Equation (9-33) where there is no possibility of confusion, we shall not bother to enclose the substrate or other species in parentheses to represent concentration [i.e., CS ≡ (S) ≡ S]. The corresponding plot in terms of substrate concentration is shown in Figure 9-7.
Figure 9-7. Evaluating Vmax and KM from batch reactor data.

Example 9-3. Batch Enzymatic Reactors
Calculate the time needed to convert 99% of the urea to ammonia and carbon dioxide in a 0.5-dm3 batch reactor. The initial concentration of urea is 0.1 mol/dm3, and the urease concentration is 0.001 g/dm3. The reaction is to be carried out isothermally at the same temperature at which the data in Table E9-2.2 were obtained.
We can use Equation (9-32),
![]()
where KM = 0.0266 mol/dm3, X = 0.99, and Curea0 = 0.1 mol/dm3, Vmax was 1.33 mol/dm3·s. However, for the conditions in the batch reactor, the enzyme concentration is only 0.001 g/dm3, compared with 5 g/dm3 in Example 9-2. Because Vmax = Et · k3, Vmax for the second enzyme concentration is

Substituting into Equation (9-32)

Analysis: This example shows a straightforward Chapter 5 type calculation of the batch reactor time to achieve a certain conversion X for an enzymatic-reaction with a Michaelis–Menten rate law. This batch reaction time is very short; consequently, a continuous flow reactor would be better suited for this reaction.
Effect of Temperature
The effect of temperature on enzymatic reactions is very complex. If the enzyme structure would remain unchanged as the temperature is increased, the rate would probably follow the Arrhenius temperature dependence. However, as the temperature increases, the enzyme can unfold and/or become denatured and lose its catalytic activity. Consequently, as the temperature increases, the reaction rate, –rS, increases up to a maximum and then decreases as the temperature is increased further. The descending part of this curve is called temperature inactivation or thermal denaturizing.10 Figure 9-8 shows an example of this optimum in enzyme activity.11
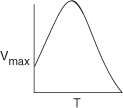
Figure 9-8. Catalytic breakdown rate of H2O2 depending on temperature.

Courtesy of S. Aiba, A. E. Humphrey, and N. F. Mills, Biochemical Engineering, Academic Press (1973).
Side Note: Lab-on-a-Chip. Enzyme-catalyzed polymerization of nucleotides is a key step in DNA identification. The microfluidic device shown in Figure SN9.1 is used to identify DNA strands and was developed at the University of Michigan.
Figure SN9.1. Microfluidic device to identify DNA.

Courtesy of Science, 282, 484 (1998).
In order to identify the DNA, its concentration must be raised to a level that can be easily quantified. This increase is typically accomplished by replicating the DNA in the following manner. After a biological sample (e.g., purified saliva, blood) is injected into the microdevice, it is heated, and the hydrogen bonds connecting the DNA strands are broken. After breaking, a primer attaches to the DNA to form a DNA primer complex, DNA*. An enzyme ![]() then attaches to this pair, forming the DNA* enzyme complex, DNA* • E. Once this complex is formed a polymerization reaction occurs as nucleotides (dNTPs—dATP, dGTP, dCTP, and dTTP—N) attach to the primer one molecule at a time, as shown in Figure SN9.2. The enzyme interacts with the DNA strand to add the proper nucleotide in the proper order. The addition continues as the enzyme moves down the strand attaching the nucleotides until the other end of the DNA strand is reached. At this point the enzyme drops off the strand and a duplicate, double-stranded DNA molecule is formed. The reaction sequence is
then attaches to this pair, forming the DNA* enzyme complex, DNA* • E. Once this complex is formed a polymerization reaction occurs as nucleotides (dNTPs—dATP, dGTP, dCTP, and dTTP—N) attach to the primer one molecule at a time, as shown in Figure SN9.2. The enzyme interacts with the DNA strand to add the proper nucleotide in the proper order. The addition continues as the enzyme moves down the strand attaching the nucleotides until the other end of the DNA strand is reached. At this point the enzyme drops off the strand and a duplicate, double-stranded DNA molecule is formed. The reaction sequence is
Figure SN9.2. Replication sequence.
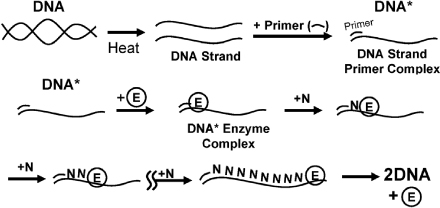
The schematic in Figure SN9.2 can be written in terms of single-step reactions where N is one of the four nucleotides.
Complex formation:
![]()
Nucleotide addition/polymerization
DNA* · E + N → DNA* · N1 · E
DNA* · N1 · E + N → DNA* · N2 · E
The process then continues much like a zipper as the enzyme moves along the strand to add more nucleotides to extend the primer. The addition of the last nucleotide is
DNA* · Ni–1 · E + N → DNA* · Ni · E
where i is the number of nucleotide molecules on the original DNA minus the nucleotides in the primer. Once a complete double-stranded DNA is formed, the polymerization stops, the enzyme drops off, and separation occurs.
Here two DNA strands really represents one double-stranded DNA helix. Once replicated in the device, the length of the DNA molecules can be analyzed by electrophoresis to indicate relevant genetic information.
9.3 Inhibition of Enzyme Reactions
In addition to temperature and solution pH, another factor that greatly influences the rates of enzyme-catalyzed reactions is the presence of an inhibitor. Inhibitors are species that interact with enzymes and render the enzyme ineffective to catalyze its specific reaction. The most dramatic consequences of enzyme inhibition are found in living organisms, where the inhibition of any particular enzyme involved in a primary metabolic pathway will render the entire pathway inoperative, resulting in either serious damage or death of the organism. For example, the inhibition of a single enzyme, cytochrome oxidase, by cyanide will cause the aerobic oxidation process to stop; death occurs in a very few minutes. There are also beneficial inhibitors, such as the ones used in the treatment of leukemia and other neoplastic diseases. Aspirin inhibits the enzyme that catalyzes the synthesis of the module prostaglandin, which is involved in the pain-producing process. Recently the discovery of DDP-4 enzyme inhibitor Januvia has been approved for the treatment of Type 2 diabetes, a disease affecting 240 million people worldwide (see P9-14B).
The three most common types of reversible inhibition occurring in enzymatic reactions are competitive, uncompetitive, and noncompetitive. The enzyme molecule is analogous to a heterogeneous catalytic surface in that it contains active sites. When competitive inhibition occurs, the substrate and inhibitor are usually similar molecules that compete for the same site on the enzyme. Uncompetitive inhibition occurs when the inhibitor deactivates the enzyme-substrate complex, sometimes by attaching itself to both the substrate and enzyme molecules of the complex. Noncompetitive inhibition occurs with enzymes containing at least two different types of sites. The substrate attaches only to one type of site, and the inhibitor attaches only to the other to render the enzyme inactive.