
9
Media Message Processing and the Embodied Mind
Measuring Bodily Responses to Open the Black Box
ABSTRACT
This chapter critically reviews research which uses physiological and behavioral measures as covert, real-time measures of motivated cognitive processing. It is written from the common perspective in psychophysiological research that the mind is an embodied organ and therefore is sustained by the body. As a result, things which occur in the mind can be “seen” in the body. Thinking, feeling, and being are all dependent on the continuous electrical, chemical, and biological functioning of the brain. As the amount of thinking and feeling varies, so do the metabolic demands of the brain. As those metabolic demands vary, so does the autonomic functioning of the body. Careful measurement of bodily responses in controlled contexts can yield fine-grained information about thinking and feeling. Recent research has applied these measures to the problem of understanding media psychology. This chapter will discuss how variations in media structure and content influence motivational activation, which in turn influences emotional responses and cognition, and how psycho-physiological and behavioral measures can be used to track the real-time processing of mediated messages.
Mind, Body, and Media
This chapter takes the perspective that the embodied mind meets mediated messages in specific local environments embedded in culturally imbued social systems. The perspective is one of systems within systems; the cognitive system is embedded in a motivational system, which is embedded in a biological system, which is embedded in a social system, which is embedded in the cultural system. Further, it is assumed that all these systems are continuously interacting with one another over time creating one large and many smaller dynamic systems. Within this perspective, the task of the communication scholar is to understand the interactions between and within one or more of these systems (Lang & Ewoldsen, 2010). The chapter focuses on the interactions between the cognitive and motivational systems within the biological system. This is not to say that the interactions of these three systems with the local environmental system or with the larger social and cultural system are not important or interesting. It is simply to admit that focusing merely on these three systems is sufficiently complex for a start.
In many ways, this perspective is antithetical to traditional approaches to studying mediated messages (Lang, 2011). First, it eschews traditional distinctions not only between media but also between mediated and interpersonal communication. From this perspective all stimuli are messages and all messages are mediated by the sensory, cognitive, motivational, and biological systems in which a conscious mind emerges. Second, it denies the existence of stability and of effects and puts in their place the concept of continuous dynamic change in which certain patterns of interaction among systems are more stable and more likely while other patterns of interaction among systems are less stable and less likely. From this perspective, a purposive strategic mediated message propagates through the systems and through time causing variation in the interactions within and among the systems both in real time (what one might call the online processing of, or interaction with, the message) and, following the conclusion of the message, in some kind of internal system time where information gleaned from the message becomes part of the system and influences future interactions among all the systems. Thus, information (in a very catholic sense) which has been, perhaps purposefully, perhaps not, placed in a message (by the world, by a producer, or by a single human being) causes immediate and somewhat predictable real-time variation in the sensory, motivational, and biological systems which encounter it, whose function is to support the perception, encoding, and storage of an idiosyncratic mental representation of that information. Introduction of this information into the system thereby changes the system and all its future behaviors, though the extent of those changes is certainly dependent upon the nature of both the information and the message that delivered it. This perspective places communication at the intersection of the biological/motivational/cognitive system and the message.
Truly accepting a dynamic systems perspective makes the actual time during which the interaction between a person and a message is occurring of primary importance. Given that we can control the structure, the timing, and the content of the message we have an opportunity, as researchers, to “observe” how the system responds to the introduction of the message. To the extent that we can observe each of the nested systems (motivational, cognitive, biological) interacting with and responding to the message, we can come to understand, explain, and to some extent predict and potentially change the system. Time is of primary importance because change within the message from one moment to the next results in subsequent change within the tripartite human system from one moment to the next, which alters how the system interacts in the next moment with the changing message. Research from this perspective can eventually learn how variation in a myriad of structural and content features of messages can drive the motivated cognitive system towards different trajectories of interaction which may be desirable.
Studying communication this way requires the ability to track changes in messages and changes in the motivational, biological, and cognitive systems in real time and the development of theories, methods, and strategies for examining the interactions among them. In this way, we can begin to understand how certain types of content or structural features in messages predictably influence dynamic change in the biological motivated cognitive system that is a message recipient. To do so requires dynamic measures of both change in messages and change in the human-motivated cognitive system.
Given that this perspective is based in part on currently popular and historically venerable non-Cartesian traditions in psychology, epitomized by psychophysiology (Cacioppo et al., 2000), which views the brain as an embodied organ inextricably linked in to bodily functions which both support and reflect its activity, physiological measures seem a natural tool to use to provide a window into the operations of the motivated cognitive system. Psychophysiology is the science of linking physiological measures with psychological correlates (Caccioppo, Tassinary, & Bernston, 2000). It is important to fundamentally understand that physiological measures are indeed measures of variation in physiological systems. That is they are measurably precise veridical (exact and actual) measures of the operation of some specific aspect of physiology such as voltage potential in a muscle, electrical conductance across a variably moist surface, or the opacity to infrared light of a portion of the body. And they measure these within some specific and known margin of error both for the measure and across time. The measurement task is merely engineering. The science is in the work that has been done over the last hundred or so years to determine, in specific environments, how, in response to a controlled stimulus, these aspects of the biological system that we can measure correlate with theorized motivational, emotional, cognitive, and behavioral responses to that stimulus – hence the term psychophysiology.
In other words, physiological measures do not provide an actual specific record of specific psychological states. Rather, in known circumstances with understood stimuli, they are indicators of psychological states. Research in psychophysiology has demonstrated large and robust correlations between specific stimuli and specific physiological responses in specific controlled environments. Over the last 30 years a fair amount of work has been done to translate that work into the communication laboratory (Lang, 1994a). This has entailed enlarging the range of stimuli from simple tones, words, and images to dynamic audio and visual stimuli of all sorts and ascertaining whether these known large and robust correlations between stimuli and physiological measures hold in the more chaotic environment of the media message. In addition, while change over time is an essential part of the psychophysiological tradition, work with media has necessarily enlarged the timeframe from milliseconds and seconds to minutes and hours. Thus, importing psychophysiological methods and theory into communication research has entailed an increase in the complexity of the stimuli and of the measurement and a concomitant decrease in control over many aspects of both the stimulus and the response. Despite these difficulties, it has also provided us with the ability to observe and learn from real-time interactions between messages and the motivated cognitive system.
As may be clear from this introduction, both the observation and interpretation of the measures are necessarily theory driven. While a statement about an increase in activation in a facial muscle might be thought of as a simple observational statement, taking the next step, and claiming that it represents a change in an emotional or psychological state requires a theory of cognition, a theory of emotion, and a theory linking the stimulus properties of the message to cognition and emotion. Much of the research discussed in this chapter and much of the importation of various physiological measures into communication research has been done either in conjunction with the development of or under the auspices of the Limited Capacity Model of Motivated Mediated Message Processing or LC4MP (Lang, 2000, Lang, 2006a, 2006b). This model draws heavily on theoretical programs in psychology (Kahneman, 1973; Shiffrin & Schneider, 1977), psychophysiology (Cacioppo & Sandman, 1981; Greenwald, Hamm, Bradley, & Lang, 1988; Lang, Simons, & Balaban, 1997b), and communication research (Bryant, Carveth, & Brown, 1981; Zillmann, 1982a). Its primary pieces were developed by others and integrated and applied to communication in an attempt to explain the real-time dynamic interaction between messages and the human biological, motivated, cognitive system. This chapter lays out the LC4MP, the embedded use of psychophysiological measures in communication, and discusses potential developments in this area.
The Theoretical Model
The LCModelMMMP presents a plausible explanation for how information moves from the message to the mind. The model is itself dynamic, minimalist and data-driven and seeks to remain compatible with our ever-increasing understanding of neuro- bio- cognitive- psychology. As a result of these characteristics, the model incorporates the fewest possible theoretical variables and concepts needed to accomplish its task and strives to develop, for each concept, at least one specific measurement. Thus, unlike traditional effects research, where something about a message is manipulated and something about a person is measured after the message has been consumed, and these pre-and post-data are used as supporting evidence for what occurred in between; LC4MP attempts to measure every hypothesized dynamic interaction between the message and the motivated cognitive system in real time.
The LCMMMMessageProcessing takes the perspective that humans have a cognitive system capable of performing at least three specific processes, embedded in a motivational system designed by evolution to promote survival, embedded in the physiological and biological system which is the body. Variations in any of these systems caused by external (as in a mediated message or a conversational partner) or internal (as in a thought or an upset stomach) stimuli have immediate impact on the functioning of the cognitive system. The three specified processes in the cognitive system are encoding, storage, and retrieval.
Encoding is the idiosyncratic non-veridical process by which information contained in the external world (e.g., the mediated message) is turned into a mental representation. There are three important things to understand about this process. First, it occurs over time. This means that the amount of time the system has to encode a message significantly impacts the amount of information that is incorporated into the resulting mental representation. Second, encoding is non-veridical: Eyes are not cameras and ears are not audio recorders. Rather, seeing and hearing are driven by both hard-and software. They are constructive processes, which rely on the external information in the world and internal information which has been learned over a lifetime of interaction within and with the world. Thus, encoding produces not an exact representation of reality but rather a constructed one. Further, the construction rests not only on information from the external world but also on the experiences of the encoder. Thus, each person's representation is idiosyncratic. The aspects of the external stimulus selected for encoding will vary based on the interests, knowledge, and background of the person doing the encoding. This is not to say that each person will encode a completely unrelated representation of a given stimulus because the evolutionary mechanical characteristics of the biological system will ensure the encoding of aspects of stimuli directly related to survival. Similarly, psychological characteristics of stimuli will elicit from all encoders various automatic and reflexive cognitive and motivational responses, which ensure their inclusion in the final representation. Hence, the core of encoded representations, those aspects related to the basic functioning of the motivated cognitive system, will be there for everyone. What will vary between people is the selection from the myriad of other details which accompany that basic psychological core. Thus, when a fast-moving car comes screeching around the corner, careers off a lamp post, and rockets out of sight down an alley, all observers will report accurately that it was a vehicle, that it was fast-moving, the general direction from which it came, and more accurately the direction in which it disappeared. Other details will depend on the observers. Car buffs may report on make and model, an artist might notice the color, the off-duty policeman may come up with a license plate number, and inveterate people watchers may notice something about the driver. These people will be selecting the details that are associated with how they have learned to see the world.
Storage is the process by which encoded mental representations become part of what a human knows. The LC4MP considers memory to be essentially neurologically based. It is likely that what a person is thinking about at any given time corresponds to the set of activated neurons which make up the representations of those things. Neurons which fire together, wire together and likely form in some way the neurological basis underlying association models of memory. The LC4MP does not endorse a specific model of memory but rather assumes that simultaneously active memories become linked and that activation can spread through the memory network. Thus, encoding creates an active mental representation of what is occurring in real time. Aspects of the external stimuli which are related to or match information already known activate their stored mental representations during this process and the simultaneous activation links the new information to the old. The process is largely automatic though people can work to increase the efficiency of the storage process.
Retrieval is the process of activating previously stored information. It happens automatically in response to stimuli in the external world (e.g., when you see an elephant, you effortlessly retrieve the label elephant and the knowledge that it is a grey animal with a trunk, etc.) and it can happen under the control of a person trying to remember what they know (e.g., can I remember if it is Indian or African elephants that have big ears?).
The LimitedCapacity4MP also argues that people do not have an unlimited ability to take in, store, or retrieve information. The model conceives of humans as limited capacity processors and conceptualizes attention as the allocation of this limited capacity of processing resources (Grimes & Meadowcroft, 1995; Kahneman, 1973; Lynn, 1966; Ohman, 1979; Reeves et al., 1985; Wickens, 1984). As a result, what is encoded, what is stored, and what can be retrieved is only a subset of the information contained in the world or in a mediated message. Many aspects of the environment, the processing system, and the message directly influence how these capacity limitations impact our ability to perceive, comprehend, and remember the information contained in messages. Among the most important external variables are the amount of information, the rate of information presentation, and the motivational relevance of the information. Among the most important internal variables are practice, prior knowledge, and expertise.
The model theorizes that limited cognitive resources are allocated independently (through both automatic and controlled mechanisms) to encoding, storage, and retrieval. Aspects of the environment which represent change, often associated with novelty (e.g., someone opening a door or the sound of a car starting) or that are learned signals of import (e.g., your name or a siren) elicit orienting responses (ORs) and thereby the automatic allocation of resources to encoding. Aspects of the environment that are related to threats and opportunities automatically elicit motivational activation and thereby the allocation of resources to encoding and storage. When more resources are required to process a stimulus than have been allocated, processing performance is impaired and a form of cognitive overload occurs.
Within the LCMMMediatedMP encoding, storage, and retrieval, though capable of being influenced by intent, generally operate automatically and outside conscious awareness and control. Further, these are primitive, built-in, biologically based cognitive mechanisms developed by evolution and not subject to change within the relatively short time encompassed by human history. For this reason, a primary assumption of the LC4MP is that these processes initially respond to mediated stimuli as if they were real (Reeves & Nass, 1996). Certainly, as people interact with mediated stimuli their conscious knowledge that they are not in fact precisely “real” does function to influence these responses and alter how the system is operating. However, they cannot stop these processes from operating, they can only inhibit, shift, or facilitate them.
Finally, the LCMotivatedMMMP theorizes that the cognitive system is embedded within a biological motivational system that was designed by evolution to promote survival of the species and the individual. The motivational system consists of two independently operating systems, the appetitive or approach system and the aversive or avoid system (Cacioppo & Gardner, 1999). Activation in these systems varies continuously across time in response to external and internal stimuli representing threats and opportunities. The appetitive system activates in response to opportunities while the aversive system activates in response to threat. At any given time, the environment may contain threats, opportunities, both or neither. For this reason, the systems' activation levels vary independently and automatically as needed. The two systems have somewhat different operating characteristics. Neither system is ever off; however there is a very low level of activation, which we might think of as idling. Dual systems theory suggests that in a neutral environment (e.g., one with no threats and no opportunities) both the appetitive and the aversive system will be idling, and, when that occurs, the activation in the appetitive system is greater than that in the aversive system. The difference between these two is called the positivity offset and its theorized purpose is to ensure that people will venture out into the world to search for opportunities. To make up for this lower level of idling activation, the aversive system is thought to react more quickly and more vigorously to potential threats. This characteristic is called the negativity bias. Its primary purpose is to ensure survival through swift threat avoidance, it being best to avoid probable threats first and think about it later rather than to fail to avoid and thereby think no more.
In addition to motivating behavior (e.g., approach and avoidance) the motivational systems are also theorized to motivate cognition (Lang, Bradley, & Cuthbert, 1997a). That is, at the same time that motivational activation may be driving increased blood flow to the muscles or the brain to support running or thinking; it is also fine tuning the distribution of finite cognitive resources among the processes involved in thinking about the world (e.g., encoding, storage, and retrieval). Appetitive or approach activation is theorized to be strongly associated with information intake as information increases our ability to take advantage of opportunities. Thus the model predicts that as appetitive activation increases more cognitive resources are allocated to encoding and storage external information. On the other hand, aversive activation is primarily related to avoiding threat. Therefore, at low to moderate levels of aversive activation, where there are only potential, not imminent, threats, resources are allocated to scanning the environment and taking in information. However, once a threat has been identified, as danger becomes more immediate, resources are withdrawn from the task of taking in additional information about the threat, and are allocated instead to the internal tasks of retrieving information about how to respond in the current environment, and towards making decisions about what to do (Yegiyan & Lang, 2010).
In the context of communication research, this means that messages which contain motivationally relevant content automatically activate the motivational systems resulting in a change in how the cognitive system distributes its finite resources and therefore a change in the amount of information which is encoded, stored, and retrieved. Motivationally relevant content includes any content that represents threats or opportunities. Some motivationally relevant content is determined by evolution, these things represent hardwired threats and opportunities. Hardwired opportunities include food and sex. Hardwired threats include signs of distress (screaming, weeping), fast onset stimuli, and intense (e.g., very loud, very bright, very smelly) stimuli. Thus, messages which contain strong emotion, food, naked people, high-intensity fast-moving action, snakes, spiders, and so on, are likely to elicit automatic motivational activation in most users. In addition, people learn through experience to associate motivational activation with a myriad of objects, words, expressions, places, and things. Some of these associations are cultural, some are specific to a given social system, and some are purely personal. Once learned, however, these things also automatically elicit motivational activation.
The Operational Model
As stressed above, all of this theorizing is not worth much if there are no measures associated with the various parts of the model. A set of cognitive and physiological time-based measures shown to be correlated with the motivational and cognitive processes discussed above have been developed, borrowed, and adapted for use in the communication laboratory. Many of these measures are familiar to communication researchers. However, because of the dynamic nature of the theory they may be employed somewhat differently when used from this theoretical perspective. For example, rather than looking at the average of the measure over an entire message, these measures are tracked over the time-course of the message and the over-time interactions between change in the message and change in the motivated cognitive system are assessed. This allows for the measure of real-time, dynamic processing of the message.
In addition, research from this perspective requires dynamic measures of psychologically and motivationally relevant aspects of mediated messages since it is necessary to track the variation in signal, novel, and motivationally relevant information within the message as well as the change in variables related to the level of resources required to successfully encode and store the information contained in the message (Detenber & Lang, 2011). The next section will first present the motivated cognitive measures and then the dynamic message measures.
Motivated Cognitive Measures
The model as presented above makes predictions about motivational activation, orienting responses, resource allocation, encoding, storage, and retrieval. Therefore it requires dynamic measures of appetitive and aversive activation, encoding, storage, and retrieval, and the allocation of finite processing resources.
Encoding, storage, and retrieval performance are indexed by recognition, cued recall, and retrieval, respectively (Lang, 2000; Zechmeister & Nyberg, 1982).
Recognition is measured by presenting participants with an exact representation of what they saw or heard and asking them to decide whether or not they saw it. When measuring recognition it is not necessary for the participant to locate or retrieve information, they need only to perceive the stimulus and decide if they have seen it before. This is an appropriate measure of encoding, because people encode a great deal more information than they actually successfully store or can retrieve, but the process of encoding information leaves behind a residue of familiarity that allows people to determine, with remarkable accuracy, whether something has been seen before. Thus, accuracy on recognition tests is a good indicator of encoding performance. In addition to accuracy, signal detection analysis can also be used which takes into account the decision-making process involved in giving a “yes/no” answer and results in two measures, D prime which is an indicator of encoding performance and criterion bias which is an indicator of the level of familiarity required by the participant in order to give a “yes” answer (Fox, Park, & Lang, 2007; Shapiro, 1994). For example, a person has a certain sense of familiarity about a recognition target – if there is a reward for getting the right answer and no punishment for getting it wrong, people will have a bias towards saying “yes I saw that before” at a lower level of familiarity. Obviously, if being wrong is punished, they will have the opposite bias.
Cued recall measures are used as an indicator of storage. Cued recall procedures require that the participant be provided with a specific cue to the location of the detail you wish them to remember. In this case, the participant does not have to find where in-memory information is located, but needs to provide only the detail. Fill in the blank questions can be used to create time-locked cued recall tests.
Finally, retrieval can be indexed using free recall measures; people are asked to recall everything they can about a stimulus. This information can be coded as to when it occurred in the message providing an over-time retrieval measure. However, this technique only assesses retrieval of a message after it has been seen. The model also posits that during message processing people engage in concurrent retrieval. That is, they are calling up information from memory that is relevant to processing the current information. At this time, the model does not have a measure of concurrent retrieval.
The allocation of processing resources is assessed in a number of ways. First, secondary task reaction times (STRTs) have been used as an indicator of resource allocation (Lang & Basil, 1998; Lang, Bradley, Park, Shin, & Chung, 2006). In this paradigm, participants carry out a specific task (e.g., watching a mediated message). Periodically, a signal occurs to which they respond, usually by pushing a button, as quickly as possible. The speed with which participants respond to the periodic signal indicates the level of resources allocated to encoding the mediated message but not required for that task, called available resources. Message processing, in particular encoding, is thought to be highest when available resources are high and lowest when available resources are low or nonexistent. Structural and content features of the message contribute both to the number of resources allocated to processing the message (as a result of signal, novel, and motivationally relevant content which automatically elicit resource allocation) and the number of resources required to process the message (as a result of complexity and difficulty). Thus, the level of available resources is dynamically changing over the course of the message in response to these content and structural features. Probes are placed periodically and strategically throughout the message in order to test predictions about level of available resources as a function of message structure and content.
Within the LC4MP, variation in resource allocation during media use is also indexed by heart rate (Lang, 1994b). Parasympathetic innervations of the heart result in heart rate deceleration during periods of intense attention to external stimuli. Heart rate can be measured time-locked to media messages and analyzed in such a way as to provide information on both short-term, called phasic, and long-term, called tonic, variations in heart rate thought to be correlated with short-and long-term changes in resource allocation. Heart rate provides a more continuous and dynamic measure than STRT.
Messages contain both structural and content features, which introduce signal or novel content and therefore elicit orienting responses. Orienting responses result in a short-term increase in resources allocated to processing incoming stimuli and are indexed by a phasic (5 to 7 seconds) slowdown in heart rate. This short-term heart rate deceleration can be measured at specific points in messages where orienting is thought to occur. Over longer periods of time, sustained long-term deceleration in heart rate has been shown to be correlated with increased cognitive effort (defined in the LC4MP as total resources allocated to processing the message). Thus, by looking at the overall change in the trend of a heart rate series and the phasic variation in heart rate around that trend, one can index both short-and long-term changes in resource allocation.
Motivational activation is assessed using a set of physiological correlates of motivational activation developed in research done to explicate dimensional theories of emotion (Bradley & Lang, 2000; Lang, Gilmore, Cuthbert, & Bradley, 1996). Within these theories, emotional experience is mapped onto two primary dimensions, arousal (ranging from calm to excited), and hedonic valence (ranging from unpleasant to pleasant). These theories posit that emotion arises as a result of underlying activation in the appetitive and aversive motivational systems. The direction of motivational activation, to approach or to avoid, maps onto the hedonic valence dimension. The extent of activation maps onto the arousal dimension. Research from this perspective has shown that aversive and appetitive activation are correlated with self-reported unpleasant and pleasant emotion, respectively. In addition several physiological measures including facial electromyography (EMG) (Bolls, Potter, & Lang, 2001), skin conductance (SC) (Lang, Bolls, Potter, & Kawahara, 1999), startle, and post-auricular response (PA response) (Sparks & Lang, 2010) are correlated with activation in the motivational systems.
EMG measures electrical activity in the facial musculature. Activation in three primary facial muscle groups has been shown to be correlated with emotional experience and, presumably, the underlying motivational activation. Measurable variation in the electrical activity of these muscles occurs long before there is visual change in facial expression. Activation in the corrugator muscle group, located to the inside of the eyebrows and involved in frowning, is correlated with negative emotional experience and aversive activation (Bolls & Lang, 2003). Activation in both the zygomatic major muscle group (located on either side of the mouth and involved in smiling) and in the orbicularis oculi muscle (which rings the eye) is correlated with positive emotional experience and appetitive activation. Facial EMG measures can be collected time-locked to media use allowing the assessment of moment-by-moment change in response to real-time change in messages.
Startle and PA responses are probe measures thought to reflect activation in the aversive and appetitive systems. The startle response is a primarily aversive response which is elicited by zero-rise time (i.e., stimuli which reach their maximum level almost instantaneously) or extremely intense (e.g., loud, bright, etc.) stimuli (Lang, Bradley, & Cuthbert, 1990). Startle responses vary from quite small to what is called the whole body startle. Their primary purpose is to interrupt ongoing processing, force the processing of a potential threat, and physically protect the body. An obligatory eyeblink is one component of the startle response. In the laboratory, startles are elicited by a very brief no rise-time burst of white noise. Electrodes placed on the orbicularis oculi muscle are used to measure the contraction of the muscle, which results in the eyeblink. The size of the blink is correlated with aversive and appetitive activation. At low levels of aversive activation startles are small, at high levels of aversive activation they are large. At low levels of appetitive activation startles are small and at high levels of appetitive activation they are even smaller. Aversive activation is thought to facilitate startle while appetitive activation inhibits it.
The PA response is thought to have been correlated primarily with appetitive activation (Sparks & Lang, 2010). The electrodes placed on the post-auricular muscle, located behind the ears, measure contractions elicited by the same white noise probe that elicits the startle. PA responses are small during aversive activation or low levels of appetitive activation but much larger during high levels of appetitive activation.
Both PA and startle responses are probe measures, like STRT, which means they can be placed at strategic points in a mediated message to provide information about appetitive and aversive activation at a specific moment in time. Multiple probes placed over the course of the message provide insight into variation over longer periods of time.
The arousal dimension of emotion, thought to be indicative of activation in the appetitive and/or aversive motivational systems, is correlated with skin conductance (SC). SC is measured by running a very slight DC current between electrodes placed on the palm. Increases and decreases in the sympathetic division of the autonomic nervous system, correlated with the experience of arousal, result in the raising and lowering of the level of sweat in the eccrine sweat glands. Thus, with increasing sympathetic arousal, the level of sweat in the glands rises, resulting in faster conductance of the current across the surface of the hand, and thereby increased skin conductance.
Heart rate, skin conductance, facial EMG, startle, PA response, and STRT can all be measured time-locked to use of a given medium providing a picture of moment-by-moment change in appetitive and aversive activation and resource allocation. After media use, memory measures which track recognition, cued recall, and free recall for information occurring at specific time points over the course of a message, can provide us with an over-time assessment of cognitive processing. This information can then be used to assess the dynamic interactions between the medium and the motivated cognitive system. Future research in this area should continue to search for more and better indicators of the important processes and mechanisms posited by the model. Indeed some recent research has begun looking at functional magnetic resonance imaging (fMRI) as an indicator of motivational activation during risky behavior (Potter, Lang, Brown, Fukunaga, & Krawitz, 2008b).
Dynamic Message Measurements
This brings us to the problem of how to measure dynamic change in psychologically relevant aspects (that is those that are thought to influence the motivated cognitive system) of mediated messages. LC4MP is primarily concerned with three types of psychologically relevant message information: (1) those aspects of a message which elicit orienting (e.g., signal and novel stimuli or structures), (2) the amount of information in the message requiring resources in order to be processed (e.g., complexity, difficulty), and (3) motivationally relevant information (e.g., speed, rise time, pleasant and unpleasant information).
Because LC4MP theorizes that novel and signal aspects of mediated messages will elicit orienting responses which result in an increase in resources allocated to processing, we need to know which aspects of each medium function as novel and signal stimuli and to have a dynamic measure of these elements. The first step in developing these measures is to determine which aspects of any given medium elicit orienting. To date, research has developed a list of both content and structural elements in several media that have been shown to elicit orienting responses, these elements are called orienting eliciting structural features or OESFs. For example, in TV, camera changes, movement towards the camera, certain changes in form, and some forms of camera movement have been shown to elicit orienting responses (Lang, 1990). In web-based media, the appearance of emotional pictures, warnings, and animation have been shown to elicit orienting (Lang, Borse, Wise, & David, 2002). In audio messages, sound effects, emotional words, and voice changes have been shown to elicit orienting (Potter, Lang, & Bolls, 2008a). Given the list of OESFs for a medium, any media interaction can be coded over time for their occurrence. Each OESF represents a point in time when additional resources are allocated to encoding the message. Thus, the more OESFs there are, the more resources are automatically allocated to processing the message. In addition, there should be an increase in resources allocated immediately following each OESF.
Of course, while, in general, having more resources allocated should increase message processing, the actual relationship is also dependent on the number of resources required to process the information immediately following the OESF. When messages are more difficult or contain more information they require more resources to be successfully encoded. Therefore, a second measure, called information introduced (ii) is paired with the OESF measure (Lang et al., 2006; Lang, Park, Sanders-Jackson, & Wilson, 2007). The information introduced measure assesses the amount of information present before and after each OESF on a number of dimensions (five for audio information and seven for video information). The more dimensions of information present following an OESF, the greater the resources required to process it and the fewer the available resources. Combining the information introduced (resources required to process the message) and OESF (resources allocated to processing the message) measures results in an over-time measure of predicted available resources. One such recently developed combination measure is called the dynamic audio/video redundancy and complexity measure (DAVRC) (Lee, 2009). This measure produces a ratio of OESFs and a measure of the dynamic variation of information introduced for every 4 seconds of a given message. Recent research shows that it successfully predicts variation in secondary task reaction times and encoding over time within a message.
Finally, LC4MP posits that motivationally relevant material in messages will result in the automatic activation of the motivational systems, which automatically increases and redistributes the allocation of resources to message processing. Thus, we need a dynamic measure of the motivational relevance of the information contained in messages. Research from the LC4MP perspective has taken two approaches to this problem (Lang & Ewoldsen, 2010). First, coders have been asked to identify messages, or portions of messages, as containing or not containing motivationally relevant information. However, the resultant measure is far from dynamic. A second approach is to have groups of subjects view the messages and continuously rate their emotional responses (Biocca, David, & West, 1994). Specifically, one group rates how positive and another rates how negative they feel while engaging with the message. The resultant continuous response curves provide an average rating of pleasant or unpleasant emotional experience for each second of the message. These can then be used as dynamic indicators of the amount of appetitively or aversively relevant information contained in a message (Wang, Lang, & Busemeyer, 2011).
Putting it All Together
This combination of theory, over-time message coding, and time-locked measurement of the motivated cognitive system allows us to answer questions about the real-time interaction of dynamically varying messages and the motivated cognitive processing system. For example, in a recent study, stimulus messages were selected based on the dynamics of their motivationally relevant information. Messages were chosen which began neutral and became either increasingly positive (called positive) or negative (called negative), or began neutral and simultaneously became increasingly positive and negative (called coactive). Figure 9.1 shows the relevant continuous response curves.
Participants then viewed the messages while STRT, startle, PA response, facial EMG, SC, and HR data were collected. Among the theoretical predictions were that appetitive and aversive activation should increase and decrease following similar increases and decreases in the continuous response measurement (CRM) measures of positive and negative motivationally relevant information. A research question asked what would happen during coactive messages. Figure 9.2 shows results for the corrugator muscle activation which increases during aversive and decreases during appetitive activation. At the end of the message, when the messages were highly pleasant, unpleasant, or both, this muscle was highly active during negative, inhibited during positive, and in between during coactive messages. In addition, during the first startle period, when messages were relatively unemotional, there is little variation in the startle response. However, for the second startle probe, when the messages are highly emotional, startle is facilitated for negative, and inhibited for positive messages, with coactive in between.
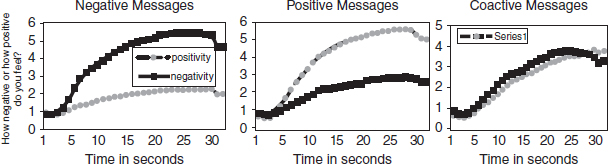
Figure 9.1 Continuous response measures of positivity and negativity.
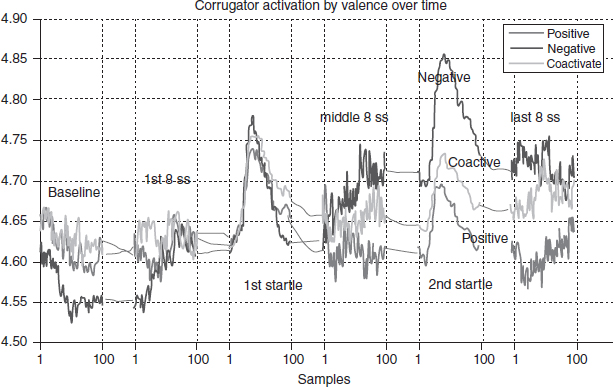
Figure 9.2 Corrugator activation.
Average measures, over time, would not have allowed the mapping of the rise in appetitive and aversive activation following emotional content. Other analyses can be done to begin to investigate the lag between message and physiological change (Yegiyan et al., 2008).
Another approach is to create models that predict physiological and behavioral responses based on continuous motivational inputs. The Dynamic Motivational Activation Model (DMA) uses continuous ratings of arousal, positivity and negativity to predict variation in skin conductance, heart rate (HR), and facial EMG (Wang et al., 2011). The model predicts between 60% and 90% of the variance in the physiological data and estimates lag times between message change and physiological change by measure.
The Future
Obviously, research from this perspective has only begun to scratch the surface of the study of communication as a fundamental human process that operates in a nested series of dynamic systems. It is worth noting here, that this model, despite its fundamentally different conceptualization of the nature of communication, does not stand in opposition to more traditional effects theories and research. Indeed, it is hoped that it can begin to shed light on the underlying mechanisms which cause some well-documented communication effects. Examples of such attempts to meld the approaches can be seen. For example, Bradley (Bradley, 2007) has shown how this approach can produce a cognitive model that displays cultivation effects. Similarly, Bucy (Bucy & Bradley, 2003, 2004) has used this approach to examine unconscious processing of political messages. It is hoped that more research will also begin to reconceptualize traditional areas of communication research and approach our field not as arbitrary beginnings, endings, and resultant effects, but rather as a continuously modifiable input into a system that can alter its patterns, resting points, and outputs. To do otherwise is to miss the beauty and complexity of this very human behavior that we study.
REFERENCES
Biocca, F., David, P., & West, M. (1994). Continuous response measurement (CRM): A computerized tool for research on the cognitive processing of communication messages. In A. Lang (Ed.), Measuring psychological responses to media messages (pp. 15–64). Hillsdale, NJ: Lawrence Erlbaum Associates.
Bolls, P., Potter, R. F., & Lang, A. (2001). The use of facial EMG to measure emotional responses to radio. Communication Research, 28, 627–651.
Bolls, P. D., & Lang, A. (2003). I saw it on the radio: The allocation of attention to high-imagery radio advertisements. Media Psychology, 5, 33–55.
Bradley, M. M., & Lang, P. (2000). Measuring emotion: Behavior, feeling, and physiology. In R. D. Lane & L. Nadel (Eds.), Cognitive neuroscience of emotion (pp. 242–276). Oxford, UK: Oxford University Press.
Bradley, S. D. (2007). Neural network simulations support heuristic processing model of cultivation effects. Media Psychology, 10, 449–469.
Bryant, J., Carveth, R. A., & Brown, D. (1981). Television viewing and anxiety: An experimental examination. Journal of Communication, 31, 106–119.
Bucy, E. P., & Bradley, S. D. (2003). Facing crisis: Attentional and EMG responses to inappropriate leader displays. Psychophysiology, 40, S30–S30.
Bucy, E. P., & Bradley, S. D. (2004). Presidential expressions and viewer emotion: Counterempathic responses to televised leader displays. Social Science Information Sur les Sciences Sociales, 43, 59–94.
Caccioppo, J. T., Tassinary, L. G., & Bernston, G. G. (2000). Psychophysiological science. In J. T. Cacioppo, L. G. Tassinary, & G. G. Bernston (Eds.), Handbook of psychophysiology (pp. 3–26). Cambridge, UK: Cambridge University Press.
Detenber, B. H., & Lang, A. (2011). The influence of media form and presentation attributes on emotion. In K. Döveling, C. V. Scheve, & E. Konjin (Eds.), Handbook of emotions in mass media (pp. 275–293). London, UK: Routledge.
Fox, J., Park, B., & Lang, A. (2007). When available resources becomes negative resources: The effects of cognitive overload on memory sensitivity and criterion bias. Communication Research, 34, 277–296.
Greenwald, M. K., Hamm, A. O., Bradley, M. M., & Lang, P. J. (1988). The acoustic startle probe in the assessment of classical aversive-conditioning. Psychophysiology, 25, 451.
Grimes, T., & Meadowcroft, J. (1995). Attention to television and some methods for its measurement. Communication Yearbook, 18, 133–161.
Kahneman, D. (1973). Attention and effort. Englewood Cliffs, NJ: Prentice Hall.
Lang, A. (1990). Involuntary attention and physiological arousal evoked by structural features and emotional content in TV commercials. Communication Research, 17, 275–299.
Lang, A. (Ed.) (1994a). Measuring psychological responses to media. Hillsdale, NJ: Lawrence Erlbaum Associates.
Lang, A. (1994b). What can the heart tell us about thinking? In A. Lang (Ed.), Measuring psychological responses to media messages (pp. 99–111). Hillsdale, NJ: Lawrence Erlbaum Associates.
Lang, A. (2000). The limited capacity model of mediated message processing. Journal of Communication, 50, 46–70.
Lang, A. (2006a). Motivated cognition (LC4MP): The influence of appetitive and aversive activation on the processing of video games. In P. Messarsis & L. Humphries (Eds.), Digital media: Transformation in human communication (pp. 237–254). New York, NY: Peter Lang.
Lang, A. (2006b). Using the limited capacity model of motivated mediated message processing (LC4MP) to design effective cancer communication messages. Journal of Communication, 56, s57–s80.
Lang, A. (2011, May). The shifting paradigm of communication research. Paper presented at the Mass Communication Division of the International Communication Association. Boston, MA.
Lang, A., & Basil, M. (1998). What do secondary task reaction times measure anyway? In M. Roloff (Ed.), Communication yearbook (Vol. 21, pp. 443–470). Beverly Hills, CA: Sage.
Lang, A., Bolls, P., Potter, R. F., & Kawahara, K. (1999). The effects of production pacing and arousing content on the information processing of television messages. Journal of Broadcasting and Electronic Media, 43(4), 451–475.
Lang, A., Borse, J., Wise, K., & David, P. (2002). Captured by the World Wide Web – orienting to structural and content features of computer-presented information. Communication Research, 29(3), 215–245.
Lang, A., Bradley, S. D., Park, B., Shin, M., & Chung, Y. (2006). Parsing the resource pie: Using STRTs to measure attention to mediated messages. Media Psychology, 8, 369–394.
Lang, A., & Ewoldsen, D. (2010). Beyond effects: Conceptualizing communication as dynamic, complex, nonlinear, and fundamental. In S. Allan (Ed.), Rethinking communication: Keywords in communication research (pp. 111–122). Cresskill, NJ: Hampton Press.
Lang, A., Park, B.-H., Sanders-Jackson, A., & Wilson, B. D. (2007). Separating emotional and cognitive load: How valence, arousing content, virtual complexity and information density affect the availability of cognitive resources. Media Psychology, 10, 317–338.
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1997a). Motivated attention: Affect, activation, and action. In P. J. Lang, R. F. Simons, & M. T. Balaban (Eds.), Attention and orienting: Sensory and motivational processes (pp. 97–135). Hillsdale, NJ: Lawrence Erlbaum Associates.
Lang, P. J., Gilmore, R., Cuthbert, B. N., & Bradley, M. M. (1996, October). Inside picture processing: Emotional modulation of ERPs from the cortical surface. Paper presented to the Society for Psychophysiological Research, October, Vancouver, BC, Canada.
Lang, P. J., Simons, R. F., & Balaban, M. T. (1997b). Attention and orienting: Sensory and motivational processes. Hillsdale, NJ: Lawrence Erlbaum Associates.
Lee, S. (2009). Understanding available resources during TV message processing: Audio/video redundancy, emotion, structural complexity, and motivated cognition. (Unpublished dissertation.) Bloomington, IN: Indiana University.
Lynn, R. (1966). Attention, arousal and the orientation reaction. Oxford, UK: Pergamon Press.
Ohman, A. (1979). The orientations response, attention, and learning: An information processing perspective. In H. D. Kimmel, E. H. V. Olst, & J. F. Orlebeke (Eds.), The orienting reflex in humans. Hillsdale, NJ: Lawrence Erlbaum Associates.
Potter, R., Lang, A., & Bolls, P. (2008a). Identifying structural features of audio: Orienting responses during radio messages and their impact on recognition. Journal of Media Psychology, 20(4), 168–177.
Potter, R. F., Lang, A., Brown, J. W., Fukunaga, R., & Krawitz, A. (2008b, May). Brain activation during risk: The influence of trait motivation on ACC activation during choice and consequence. Paper presented at the annual meeting of the International Communication Association, Montreal, QC, Canada.
Reeves, B., & Nass, C. (1996). The media equation: How people treat computers, television, and new media like real people and places. Stanford, CA: CSLI Publications.
Reeves, B., Thorson, E., Rothschild, M., McDonald, D., Hirsch, J., & Goldstein, R. (1985). Attention to television: Interstimulus effects of movement and scene changes on alpha variation over time. International Journal of Neuroscience, 25, 242–255.
Shapiro, M. A. (1994). Signal detection measures of recognition memory. In A. Lang (Ed.), Measuring psychological responses to media messages (pp. 133–148). Hillsdale, NJ: Lawrence Erlbaum Associates.
Shiffrin, R. M., & Schneider, W. (1977). Controlled and automatic human information processing: II Perceptual learning, automatic attending and a general theory. Psychological Review, 84, 127–189.
Sparks, J. V., & Lang, A. (2010). An initial examination of the post-auricular reflex as a physiological indicator of appetitive activation during television viewing. Communication Methods and Measures, 4, 311–330.
Wang, Z., Lang, A., & Busemeyer, J. (2011). Motivational processing and choice behavior during television viewing: An integrative dynamic approach. Journal of Communication, 61, 71–93.
Wickens, C. D. (1984). Processing resources in attention. In R. Parasuraman & D. R. Davies (Eds.), Varieties of attention. Orlando, FL: Academic Press.
Yegiyan, N. S., & Lang, A. (2010). Processing central and peripheral detail: How content arousal and emotional tone influence encoding. Media Psychology, 13, 77–99.
Yegiyan, N. S., Wilson, B. D., Gao, Y., Mayell, S., Wang, Z., & Lang, A. (2008, May). Approach? Avoid? Both? Processing coactive motivational media messages. Paper presented at the annual meeting of the International Communication Association, Montreal, Canada.
Zechmeister, E. B., & Nyberg, S. E. (1982). Human memory: An introduction to research and theory. Monterey, CA: Brooks/Cole.
Zillmann, D. (1982a). Television and arousal. In D. Pearl, L. Bouthilet, & J. Lazar (Eds.), Television and behavior: Ten years of scientific progress and implications for the eighties (Vol. 2). Washington, DC: US Department of Health and Human Services.
FURTHER READING
Angelini, J. R., & Bradley, S. D. (2010). Homosexual imagery in print advertisements: Attended, remembered, but disliked. Journal of Homosexuality, 57(4), 485–502.
Basil, M., Schooler, C., & Reeves, B. (1991). Positive and negative political advertising: Effectiveness of ads and perceptions of candidates. In F. Biocca (Ed.), Television and political advertising. Vol. 1: Psychological processes (pp. 245–262). Hillsdale, NJ: Lawrence Erlbaum Associates.
Bradley, S. D. (2007a). Dynamic, embodied, limited-capacity attention and memory: Modeling cognitive processing of mediated stimuli. Media Psychology, 9(1), 211–239.
Bradley, S. D., Angelini, J. R., & Lee, S. (2007). Psychophysiological and memory effects of negative political ads – Aversive, arousing, and well remembered. Journal of Advertising, 36(4), 115–127.
Bucy, E. P., Lang, A., Potter, R. F., & Grabe, M. E. (1999). Formal features of cyberspace: Relationships between web page complexity and site traffic. Journal of the American Society for Information Science, 50(13), 1246–1256.
Cacioppo, J. T., & Gardner, W. L. (1999). Emotion. Annual Reviews: Psychology, 50, 191–214.
Cacioppo, J. T., & Sandman, C. A. (1981). Psychophysiological functioning, cognitive responding and attitudes. Hillsdale, NJ: Lawrence Erlbaum Associates.
Cacioppo, J. T., Tassinary, L. G., & Berntson, G. G. (Eds.) (2000), Handbook of psychophysiology. Cambridge, UK: Cambridge University Press.
Chock, T., Fox, J., Angelini, J., Lee, S., & Lang, A. (2007). Telling me quickly: How arousing fast-paced PSAs decrease self-other differences. Communication Research, 34(6), 618–636.
Detenber, B. H., & Reeves, B. (1996). A bio-informational theory of emotion: Motion and image size effects on viewers. Journal of Communication, 46(3), 66–84.
Donohew, L., Zimmerman, R., Cupp, P. S., Novak, S., Colon, S., & Abell, R. (2000). Sensation seeking, impulsive decision-making, and risky sex: Implications for risk-taking and design of interventions. Personality and Individual Differences, 28(6), 1079–1091.
Donohew, R. L., Hoyle, R. H., Clayton, R. R., Skinner, W. F., Colon, S. E., & Rice, R. E. (1999). Sensation seeking and drug use by adolescents and their friends: Models for marijuana and alcohol. Journal of Studies on Alcohol, 60(5), 622–631.
Geiger, S. F., & Reeves, B. (1991). The effects of visual structure and content emphasis on the evaluation and memory for political candidates. In F. Biocca (Ed.), Television and political advertising. Vol. 1: Psychological processes (pp. 125–143). Hillsdale, NJ: Lawrence Erlbaum Associates.
Grabe, M. E., Zhou, S. H., Lang, A., & Bolls, P. D. (2000). Packaging television news: The effects of tabloid on information processing and evaluative responses. Journal of Broadcasting and Electronic Media, 44(4), 581–598.
Lang, A., & Basil, M. (1998). What do secondary task reaction times measure anyway? In M. Roloff (Ed.), Communication yearbook (Vol. 21, pp. 443–470). Beverly Hills, CA: Sage.
Lang, A., Bradley, S. D., Chung, Y., & Lee, S. (2003). Where the mind meets the message: Reflections on ten years of measuring psychological responses to media. Journal of Broadcasting and Electronic Media, 47(4), 650–655.
Lang, A., Chung, Y., Lee, S., Schwartz, N., & Shin, M. (2005). It's an arousing, fast-paced kind of the world: The effects of age and sensation seeking on the information processing of substance abuse PSAs. Media Psychology, 7(4), 421–454.
Lang, A., Chung, Y., Lee, S., & Zhao, X. (2005). It's the product: Do risky products compel attention and elicit arousal in media users? Health Communication, 17(3), 283–300.
Lang, A., Dhillon, K., & Dong, Q. (1995). The effects of emotional arousal and valence on television viewers' cognitive capacity and memory. Journal of Broadcasting and Electronic Media, 39(3), 313–327.
Lang, A., & Ewoldsen, D. (2011). Measuring emotion. In K. Döveling, C. von Scheve, & E. Konjin (Eds.), Handbook of emotions in mass media. London, UK: Routledge.
Lang, A., Kurita, S., Rubenking, B., & Potter, R. F. (2011). MiniMAM: Developing a short version of the Motivation Activation Measure. Communication Methods and Measures, 5, 146–162.
Lang, A., Newhagen, J., & Reeves, B. (1996). Negative video as structure: Emotion, attention, capacity, and memory. Journal of Broadcasting and Electronic Media, 40(4), 460–477.
Lang, A., Potter, R. F., & Bolls, P. D. (1999). Something for nothing: Is visual encoding automatic? Media Psychology, 1(2), 145–163.
Lang, A., Potter, R., & Bolls, P. (2009). Where psychophysiology meets the media: Taking the effects out of mass media research. In J. Bryant & M. B. Oliver (Eds.), Media effects: Advances in theory and research (pp. 185–206). New York, NY: Taylor & Francis.
Lang, A., Shin, M., Bradley, S. D., Wang, Z., Lee, S., & Potter, D. (2005). Wait! Don't turn that dial! More excitement to come! Get the effects of storyline and production pacing in local television news on channel changing behavior and information processing in a free choice environment. Journal of Broadcasting and Electronic Media, 49, 3–22.
Lang, A., Shin, M., & Lee, S. (2005). Sensation seeking, motivation, and substance use: A dual system approach. Media Psychology, 7, 1–30.
Lang, A., Sias, P. M., Chantrill, P., & Burek, J. A. (1995). Tell me a story: Narrative elaboration and memory for television. Communication Reports, 8(2), 102–110.
Lang, A., Wise, K., Lee, S., & Cai, X. (2003). The effects of sexual appeals on physiological, cognitive, emotional, and attitudinal responses for product and alcohol billboard advertising. In T. Reichert & J. Lambiase (Eds.), Sex and advertising (pp. 107–132). Mahwah, NJ: Lawrence Erlbaum Associates.
Lang, A., & Yegiyan, N. (2009). Motivated Message Processing: How media elicit motivation which influences how media are processed. In J. McCroskey, K. Floyd, & M. Beatty (Eds.), Biological dimensions of communication (pp. 135–158): Cresskill, NJ: Hampton Press.
Lang, A., Zhou, S., Schwartz, N., Bolls, P., & Potter, R. F. (2000). The effects of edits on arousal, attention, and memory for television messages: When an edit is an edit can an edit be too much? Journal of Broadcasting and Electronic Media, 44(1), 94–109.
Lee, S., & Lang, A. (2009). Discrete emotion and motivation: Relative activation in the appetitive and aversive motivational systems as a function of anger, sadness, fear, and joy during televised information campaigns. Media Psychology, 12(2), 148–170.
Leshner, G., Reeves, B., & Nass, C. (1998). Switching channels: The effects of television channels on the mental representation of television news. Journal of Broadcasting and Electronic Media, 42(Winter), 21–33.
Meadowcroft, J. M., & Reeves, B. (1989). Influence of story schema development on children's attention to television. Communication Research, 16(3), 352–374.
Newhagen, J. E., & Reeves, B. (1992). The evening's bad news: Effects of compelling negative television news images on memory. Journal of Communication, 42(2), 25–41.
Pescosolido, B. A., Martin, J. K., Lang, A., & Olafsdottir, S. (2008). Rethinking theoretical approaches to statement: A framework integrating normative influences on statement (FINIS). Social Science and Medicine, 67, 431–440.
Reeves, B. (1989). Theories about news and theories about cognition. American Behavioral Scientist, 33(2), 191–198.
Reeves, B., & Anderson, D. R. (1991). Media studies and psychology. Communication Research, 18(5), 597–600.
Reeves, B., & Geiger, S. (1994). Designing experiments that assess psychological responses to media messages. In A. Lang (Ed.), Measuring psychological responses to media messages (pp. 165–180). Hillsdale, NJ: Lawrence Erlbaum Associates.
Reeves, B., Lang, A., Thorson, E., & Rothschild, M. (1989). Emotional television scenes and hemispheric specialization. Human Communication Research, 15(4), 493–508.
Reeves, B., & Thorson, E. (1986). Watching television: Experiments on the viewing process. Communication Research, 13(3), 343–361.
Reeves, B., Thorson, E., & Schleuder, J. (1986). Attention to television: Psychological theories and chronometric measures. In J. Bryant & D. Zillmann (Eds.), Perspectives on media effects (pp. 251–279). Hillsdale, NJ: Lawrence Erlbaum Associates.
Shapiro, M. A., Lang, A., Hamilton, M., & Contractor, N. S. (2001). Information systems division: Intra-personal, meaning, attitude and social systems. In W. Gudykunst (Ed.), Communication yearbook 24 (pp. 17–49). Thousand Oaks, CA: Sage.
Stephenson, M. T., Palmgreen, P., Hoyle, R. H., Donohew, L., Lorch, E. P., & Colon, S. E. (1999). Short-term effects of an anti-marijuana media campaign targeting high sensation seeking adolescents. Journal of Applied Communication Research, 27(3), 175–195.
Thorson, E., & Lang, A. (1992). The effects of television videographics and lecture familiarity on adult cardiac orienting responses and memory. Communication Research, 19(3), 346–369.
Thorson, E., Reeves, B., & Schleuder, J. (1985). Message complexity and attention to television. Communication Research, 12, 427–454.
Thorson, E., Reeves, B., & Schleuder, J. (1986). Attention to local and global complexity in television messages. In M. L. McLaughlin (Ed.), Communication yearbook 10 (pp. 366–383). Newbury Park, CA: Sage.
Weaver, J. B., Masland, J. L., & Zillmann, D. (1984). Effects of erotica on young men's aesthetic perceptions of their female sexual partners. Perceptual and Motor Skills, 58, 929–930.
Zillmann, D. (1988). Mood management through communication choices. American Behavioral Scientist, 31(3), 327–340.
Zillmann, D. (1991a). The logic of suspense and mystery. In J. Bryant & D. Zillmann (Eds.), Responding to the screen: Reception and reaction processes (pp. 281–303). Hillsdale, NJ: Lawrence Erlbaum Associates.
Zillmann, D. (1991b). Television viewing and physiological arousal. In J. Bryant & D. Zillmann (Eds.), Responding to the screen: Reception and reaction processes (pp. 103–134). Hillsdale, NJ: Lawrence Erlbaum Associates.
Zillmann, D., & Bryant, J. (1982). Pornography, sexual callousness, and the trivialization of rape. Journal of Communication, 32(4), 10–21.
Zillmann, D., & Bryant, J. (1985). Selective exposure to communication. Hillsdale, NJ: Lawrence Erlbaum Associates.
Zillmann, D., & Bryant, J. (1987). Effects of pornography: The debate continues, a response. Journal of Communication, 37(1), 187–188.
Zillmann, D., & Bryant, J. (1994). Entertainment as media effect. In J. Bryant & D. Zillmann (Eds.), Media effects: Advances in theory and research (pp. 437–461). Hillsdale, NJ: Lawrence Erlbaum Associates.
Zuckerman, M. (1996). The psychobiological model for impulsive unsocialized sensation seeking: A comparative approach. Neuropsychobiology, 34(3), 125–129.
Zuckerman, M., & Kuhlman, D. M. (2000). Personality and risk-taking: Common biosocial factors. Journal of Personality, 68(6), 999–1029.